Citation: Fenglei Qi, Mark Mba Wright. A novel optimization approach to estimating kinetic parameters of the enzymatic hydrolysis of corn stover[J]. AIMS Energy, 2016, 4(1): 52-67. doi: 10.3934/energy.2016.1.52

| [1] | Pachauri RK, Allen M, Barros V, et al. (2014) Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. |
| [2] | MacDonald T, Yowell G, McCormack M (2001) US ethanol industry: production capacity outlook. California Energy Commission. P600-01-017. |
| [3] | Kazi FK, Fortman J, Anex R, et al. (2010) Techno-economic analysis of biochemical scenarios for production of cellulosic ethanol: Citeseer. |
| [4] |
Kazi FK, Fortman JA, Anex RP, et al. (2010) Techno-economic comparison of process technologies for biochemical ethanol production from corn stover. Fuel 89: S20-S28. doi: 10.1016/j.fuel.2010.01.001

|
| [5] |
Winchester N, Reilly JM (2015) The feasibility, costs, and environmental implications of large-scale biomass energy. Energy Economics 51: 188-203. doi: 10.1016/j.eneco.2015.06.016
|
| [6] |
Brown TR, Brown RC (2013) A review of cellulosic biofuel commercial‐scale projects in the United States. Biofuels, bioprod bioref 7: 235-245. doi: 10.1002/bbb.1387
|
| [7] |
Chiaramonti D, Prussi M, Ferrero S, et al. (2012) Review of pretreatment processes for lignocellulosic ethanol production, and development of an innovative method. Biomass Bioenerg 46: 25-35. doi: 10.1016/j.biombioe.2012.04.020
|
| [8] |
Sun Y, Cheng J (2002) Hydrolysis of lignocellulosic materials for ethanol production: a review. Bioresource technol 83: 1-11. doi: 10.1016/S0960-8524(01)00212-7
|
| [9] |
Wald S, Wilke CR, Blanch HW (1984) Kinetics of the enzymatic hydrolysis of cellulose. Biotechnol bioeng 26: 221-230. doi: 10.1002/bit.260260305
|
| [10] |
Hayes DJ (2009) An examination of biorefining processes, catalysts and challenges. Catal Today 145: 138-151. doi: 10.1016/j.cattod.2008.04.017
|
| [11] |
Sin G, Meyer AS, Gernaey KV (2010) Assessing reliability of cellulose hydrolysis models to support biofuel process design—identifiability and uncertainty analysis. Comput chem eng 34: 1385-1392. doi: 10.1016/j.compchemeng.2010.02.012
|
| [12] | Humbird D, Aden A (2009) Biochemical production of ethanol from corn stover: 2008 state of technology model: Citeseer. |
| [13] |
Morales-Rodriguez R, Meyer AS, Gernaey KV, et al. (2011) Dynamic model-based evaluation of process configurations for integrated operation of hydrolysis and co-fermentation for bioethanol production from lignocellulose. Bioresource Technol 102: 1174-1184. doi: 10.1016/j.biortech.2010.09.045
|
| [14] |
Morales-Rodriguez R, Meyer AS, Gernaey KV, et al. (2012) A framework for model-based optimization of bioprocesses under uncertainty: Lignocellulosic ethanol production case. Comput Chem Eng 42: 115-129. doi: 10.1016/j.compchemeng.2011.12.004
|
| [15] | Bansal P, Hall M, Realff MJ, et al. (2009) Modeling cellulase kinetics on lignocellulosic substrates. Biotechnol adv 27: 833-848. |
| [16] |
Kadam KL, Rydholm EC, McMillan JD (2004) Development and validation of a kinetic model for enzymatic saccharification of lignocellulosic biomass. Biotechnol prog 20: 698-705. doi: 10.1021/bp034316x
|
| [17] |
Zheng Y, Pan Z, Zhang R, et al. (2009) Kinetic modeling for enzymatic hydrolysis of pretreated creeping wild ryegrass. Biotechnol bioeng 102: 1558-1569. doi: 10.1002/bit.22197
|
| [18] |
Tsai CT, Morales-Rodriguez R, Sin G, et al. (2014) A dynamic model for cellulosic biomass hydrolysis: A comprehensive analysis and validation of hydrolysis and product inhibition mechanisms. Appl biochem biotech 172: 2815-2837. doi: 10.1007/s12010-013-0717-x
|
| [19] |
Scott F, Li M, Williams DL, et al. (2015) Corn stover semi-mechanistic enzymatic hydrolysis model with tight parameter confidence intervals for model-based process design and optimization. Bioresource technol 177: 255-265. doi: 10.1016/j.biortech.2014.11.062
|
| [20] |
Box GE, Draper NR (1965) The Bayesian estimation of common parameters from several responses. Biometrika 52: 355-365. doi: 10.1093/biomet/52.3-4.355
|
| [21] |
Stewart WE, Caracotsios M, Sørensen JP (1992) Parameter estimation from multiresponse data. AIChE J 38: 641-650. doi: 10.1002/aic.690380502
|
| [22] |
Ziegel ER, Gorman JW (1980) Kinetic modelling with multiresponse data. Technometrics 22: 139-151. doi: 10.1080/00401706.1980.10486129
|
| [23] | Conceição EL, Portugal AA (2012) Comparison of Two Robust Alternatives to the Box–Draper Determinant Criterion in Multiresponse Kinetic Parameter Estimation. Ind Eng Chem Res 51: 1118-1130. |
| [24] |
Raue A, Kreutz C, Maiwald T, et al. (2009) Structural and practical identifiability analysis of partially observed dynamical models by exploiting the profile likelihood. Bioinformatics 25: 1923-1929. doi: 10.1093/bioinformatics/btp358
|
| [25] |
Kim DW, Kim TS, Jeong YK, et al. (1992) Adsorption kinetics and behaviors of cellulase components on microcrystalline cellulose. J Ferment Bioeng 73: 461-466. doi: 10.1016/0922-338X(92)90138-K
|
| [26] |
Coello CAC, Pulido GT, Lechuga MS (2004) Handling multiple objectives with particle swarm optimization. Evolutionary Computation, IEEE Transactions on 8: 256-279. doi: 10.1109/TEVC.2004.826067
|
| [27] |
Hindmarsh AC, Brown PN, Grant KE, et al. (2005) SUNDIALS: Suite of nonlinear and differential/algebraic equation solvers. ACM T Math Software 31: 363-396. doi: 10.1145/1089014.1089020
|

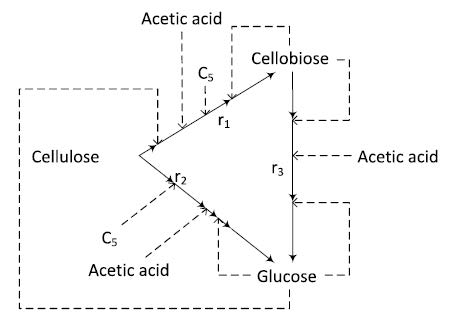
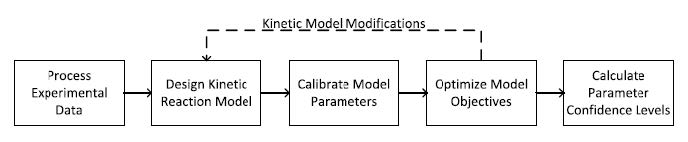
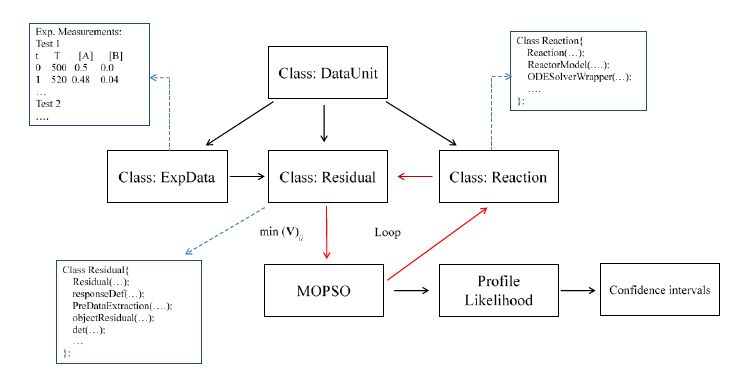
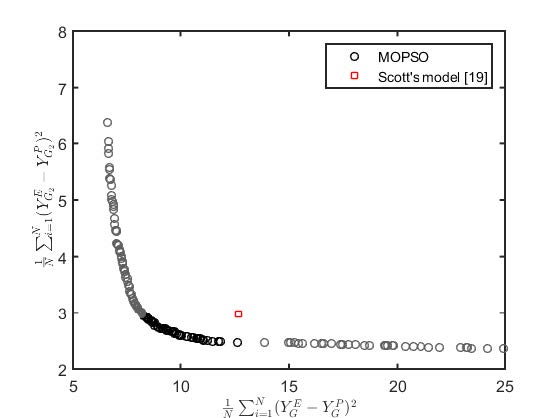
Figures(7) / Tables(3)

Fenglei Qi, Mark Mba Wright. A novel optimization approach to estimating kinetic parameters of the enzymatic hydrolysis of corn stover[J]. AIMS Energy, 2016, 4(1): 52-67. doi: 10.3934/energy.2016.1.52







 DownLoad:
DownLoad: