Citation: C. Dal Lin, M. Falanga, E. De Lauro, S. De Martino, G. Vitiello. Biochemical and biophysical mechanisms underlying the heart and the brain dialog[J]. AIMS Biophysics, 2021, 8(1): 1-33. doi: 10.3934/biophy.2021001

| [1] |
Kocica MJ, Corno AF, Carreras-Costa F, et al. (2006) The helical ventricular myocardial band: global, three-dimensional, functional architecture of the ventricular myocardium. Eur J Cardiothorac Surg 29: S21-S40. doi: 10.1016/j.ejcts.2006.03.011

|
| [2] |
Dal Lin C, Tona F, Osto E (2018) The heart as a psychoneuroendocrine and immunoregulatory organ. Sex-Specific Analysis of Cardiovascular Function 1065: 225-239. doi: 10.1007/978-3-319-77932-4_15
|
| [3] | Marinelli RA, Penney DG, Marinelli W, et al. (1991) Rotary motion in the heart and blood vessels: a review. J Appl Cardiol 6: 421-431. |
| [4] |
Sengupta PP, Narula J, Chandrashekhar Y (2014) The dynamic vortex of a beating heart: Wring out the old and ring in the new!. J Am Coll Cardiol 64: 1722-1724. doi: 10.1016/j.jacc.2014.07.975
|
| [5] | Bottio T, Buratto E, Dal Lin C, et al. (2012) Aortic valve hydrodynamics: considerations on the absence of sinuses of Valsalva. J Heart Valve Dis 21: 718-723. |
| [6] | Calgary University Medicine: Blood Moving Through the Heart - 4D Flow Available from: https://www.youtube.com/watch?v=sMeaD3Jh64E. |
| [7] |
Dal Lin C, Marinova M, Rubino G, et al. (2018) Thoughts modulate the expression of inflammatory genes and may improve the coronary blood flow in patients after a myocardial infarction. J Tradit Complement Med 8: 150-163. doi: 10.1016/j.jtcme.2017.04.011
|
| [8] |
Armour JA (2007) The little brain on the heart. Cleve Clin J Med 74: S48-S51. doi: 10.3949/ccjm.74.Suppl_1.S48
|
| [9] | Lane RD, Reiman EM, Ahern GL, et al. (1982) Activity in medial prefrontal cortex correlates with vagal component of heart rate variability during emotion. Brain Cognition 47: 97-100. |
| [10] |
Jennings JR, Sheu LK, Kuan DCH, et al. (2016) Resting state connectivity of the medial prefrontal cortex covaries with individual differences in high-frequency heart rate variability. Psychophysiology 53: 444-454. doi: 10.1111/psyp.12586
|
| [11] |
Schandry R, Montoya P (1996) Event-related brain potentials and the processing of cardiac activity. Biol Psychol 42: 75-85. doi: 10.1016/0301-0511(95)05147-3
|
| [12] |
Garfinkel SN, Barrett AB, Minati L, et al. (2013) What the heart forgets: Cardiac timing influences memory for words and is modulated by metacognition and interoceptive sensitivity. Psychophysiology 50: 505-512. doi: 10.1111/psyp.12039
|
| [13] |
Azevedo RT, Garfinkel SN, Critchley HD, et al. (2017) Cardiac afferent activity modulates the expression of racial stereotypes. Nat Commun 8: 13854. doi: 10.1038/ncomms13854
|
| [14] |
Garfinkel SN, Minati L, Gray MA, et al. (2014) Fear from the heart: Sensitivity to fear stimuli depends on individual heartbeats. J Neurosci 34: 6573-6582. doi: 10.1523/JNEUROSCI.3507-13.2014
|
| [15] |
Montoya P, Schandry R, Müller A (1993) Heartbeat evoked potentials (HEP): topography and influence of cardiac awareness and focus of attention. Electroencephalogr Clin Neurophysiol Evoked Potentials 88: 163-172. doi: 10.1016/0168-5597(93)90001-6
|
| [16] |
Thayer JF, Lane RD (2000) A model of neurovisceral integration in emotion regulation and dysregulation. J Affect Disord 61: 201-216. doi: 10.1016/S0165-0327(00)00338-4
|
| [17] |
Park G, Thayer JF (2014) From the heart to the mind: cardiac vagal tone modulates top-down and bottom-up visual perception and attention to emotional stimuli. Front Psychol 5: 278. doi: 10.3389/fpsyg.2014.00278
|
| [18] |
Thayer JF, Hansen AL, Saus-Rose E, et al. (2009) Heart rate variability, prefrontal neural function, and cognitive performance: the neurovisceral integration perspective on self-regulation, adaptation, and health. Ann Behav Med 37: 141-153. doi: 10.1007/s12160-009-9101-z
|
| [19] |
Thayer JF, Sternberg E (2006) Beyond heart rate variability: vagal regulation of allostatic systems. Ann N Y Acad Sci 1088: 361-372. doi: 10.1196/annals.1366.014
|
| [20] |
Lin PF, Lo MT, Tsao J, et al. (2014) Correlations between the signal complexity of cerebral and cardiac electrical activity: a multiscale entropy analysis. PLoS One 9: e87798. doi: 10.1371/journal.pone.0087798
|
| [21] | Aftanas LI, Brak IV, Reva NV, et al. (2013) Brain oscillations and individual variability of cardiac defense in human. Ross Fiziol Zh Im IM Sechenova 99: 1342-1356. |
| [22] |
McCraty R, Atkinson M, Bradley RT (2004) Electrophysiological evidence of intuition: Part 2. A system-wide process? J Altern Complement Med 10: 325-336. doi: 10.1089/107555304323062310
|
| [23] |
Gray MA, Beacher FD, Minati L, et al. (2012) Emotional appraisal is influenced by cardiac afferent information. Emotion 12: 180-191. doi: 10.1037/a0025083
|
| [24] |
Craig ADB (2009) How do you feel—now? The anterior insula and human awareness. Nat Rev Neurosci 10: 59-70. doi: 10.1038/nrn2555
|
| [25] |
Craig ADB (2014) How do you feel? An interoceptive moment with your neurobiological self Princeton: Princeton University Press. doi: 10.23943/princeton/9780691156767.001.0001
|
| [26] |
Grossmann I, Sahdra BK, Ciarrochi J, et al. (2016) Heart and a mind: Self-distancing facilitates the association between heart rate variability, and wise reasoning. Front Behav Neurosci 10: 68. doi: 10.3389/fnbeh.2016.00068
|
| [27] | Rahman SU, Hassan M (2013) Heart's role in the human body: A literature review. ICCSS 2: 1-6. |
| [28] | McCraty R, Trevor Bradley R, Tomasino D (2004) The resonant heart. Front Counsciousness 5: 15-19. |
| [29] | McCraty R, Atkinson M, Tomasino D, et al. (2009) The coherent heart: Heart-brain interactions, psychophysiological coherence, and the emergence of system-wide order. Integr Rev 5: 10-115. |
| [30] |
Goldstein DS (2012) Neurocardiology: therapeutic implications for cardiovascular disease. Cardiovasc Ther 30: 89-106. doi: 10.1111/j.1755-5922.2010.00244.x
|
| [31] | Dal Lin C, Poretto A, Scodro M, et al. (2015) Coronary microvascular and endothelial function regulation: Crossroads of psychoneuroendocrine immunitary signals and quantum physics. J Integr Cardiol 1: 132-163. |
| [32] |
Dal Lin C, Tona F, Osto E (2019) The crosstalk between the cardiovascular and the immune system. Vasc Biol 1: H83-H88. doi: 10.1530/VB-19-0023
|
| [33] |
Dal Lin C, Tona F, Osto E (2015) Coronary microvascular function and beyond: The crosstalk between hormones, cytokines, and neurotransmitters. Int J Endocrinol 2015: 1-17. doi: 10.1155/2015/312848
|
| [34] | Lashley KS (1942) The problem of cerebral organization in vision. Visual Mechanisms. Biological Symposia Lancaster: Jaques Cattell Press, 301-322. |
| [35] | Pribram KH (1991) Brain and Perception Hillsdale: Lawrence Erlbaum. |
| [36] | Freeman WJ (1975) Mass Action in the Nervous System New York: Academic Press. |
| [37] |
Freeman WJ (2000) Neurodynamics: An Exploration of Mesoscopic Brain Dynamics Berlin: Springer. doi: 10.1007/978-1-4471-0371-4
|
| [38] |
Ricciardi LM, Umezawa H (1967) Brain and physics of many-body problems. Kybernetik 4: 44-48. doi: 10.1007/BF00292170
|
| [39] |
Goldstone J, Salam A, Weinberg S (1962) Broken Symmetries. Phys Rev 127: 965-970. doi: 10.1103/PhysRev.127.965
|
| [40] | Umezawa H (1995) Advanced field theory: Micro, macro, and thermal physics New York: AIP. |
| [41] |
Blasone M, Jizba P, Vitiello G (2011) Quantum Field Theory and its macroscopic manifestations: Boson condensation, ordered patterns, and topological defects London: Imperial College Press. doi: 10.1142/p592
|
| [42] |
Jibu M, Yasue K (1995) Quantum brain dynamics and consciousness Amsterdam: John Benjamins Publ. doi: 10.1075/aicr.3
|
| [43] | Umezawa H (1995) Development in concepts in quantum field theory in half century. Math Jpn 41: 109-124. |
| [44] |
Vitiello G (1995) Dissipation and memory capacity in the quantum brain model. Int J Mod Phys B 9: 973-989. doi: 10.1142/S0217979295000380
|
| [45] |
Vitiello G (2001) My double unveiled Amsterdam: John Benjamins Publ. doi: 10.1075/aicr.32
|
| [46] |
Freeman WJ, Vitiello G (2006) Nonlinear brain dynamics as macroscopic manifestation of underlying many-body dynamics. Phys Life Rev 3: 93-118. doi: 10.1016/j.plrev.2006.02.001
|
| [47] |
Freeman WJ, Livi R, Obinata M (2012) Cortical phase transitions, non-equilibrium thermodynamics and the time dependent Ginzburg-Landau equation. Int J Mod Phys B 26: 1250035. doi: 10.1142/S021797921250035X
|
| [48] | Alfinito E, Vitiello G (2000) Formation and lifetime of memory domains in the dissipative quantum model of brain. Int J Mod Phys B 14: 853-868. |
| [49] |
Vitiello G (2012) Fractals, coherent states and self-similarity induced noncommutative geometry. Phys Lett A 376: 2527-2532. doi: 10.1016/j.physleta.2012.06.035
|
| [50] | Freeman W, Vitiello G (2016) Matter and mind are entangled in two streams of images guiding behavior and informing the subject through awareness. Mind Matter 14: 7-24. |
| [51] |
Del Giudice E, Doglia S, Milani M, et al. (1985) A quantum field theoretical approach to the collective behavior of biological systems. Nucl Phys B 251: 375-400. doi: 10.1016/0550-3213(85)90267-6
|
| [52] |
Del Giudice E, Doglia S, Milani M, et al. (1986) Electromagnetic field and spontaneous symmetry breakdown in biological matter. Nucl Phys B 275: 185-199. doi: 10.1016/0550-3213(86)90595-X
|
| [53] |
Del Giudice E, Preparata G, Vitiello G (1988) Water as a free electric dipole laser. Phys Rev Lett 61: 1085-1088. doi: 10.1103/PhysRevLett.61.1085
|
| [54] |
Engel GS, Calhoun TR, Read EL, et al. (2007) Evidence for wavelike energy transfer through quantum coherence in photosynthetic systems. Nature 446: 782-786. doi: 10.1038/nature05678
|
| [55] |
Peacock JA (1990) An in vitro study of the onset of turbulence in the sinus of valsalva. Circ Res 67: 448-460. doi: 10.1161/01.RES.67.2.448
|
| [56] | Mettauer B, Levy F, Richard R, et al. (2005) Exercising with a denervated heart after cardiac transplantation. Ann Transplant 10: 35-42. |
| [57] | Armour JA, Ardell JL (2004) Basic and Clinical Neurocardiology Oxford: Oxford University Press. |
| [58] |
Biasetti J, Hussain F, Gasser TC (2011) Blood flow and coherent vortices in the normal and aneurysmatic aortas: a fluid dynamical approach to intra-luminal thrombus formation. J R Soc Interface 8: 1449-1461. doi: 10.1098/rsif.2011.0041
|
| [59] |
Matsumoto H, Papastamatiou NJ, Umezawa H, et al. (1975) Dynamical rearrangement in the Anderson-Higgs-Kibble mechanism. Nucl Phys B 97: 61-89. doi: 10.1016/0550-3213(75)90215-1
|
| [60] |
Matsumoto H, Papastamatiou NJ, Umezawa H (1975) The boson transformation and the vortex solution. Nucl Phys B 97: 90-124. doi: 10.1016/0550-3213(75)90216-3
|
| [61] |
Manka R, Vitiello G (1990) Topological solitons and temperature effects in gauge field theory. Ann Phys 199: 61-83. doi: 10.1016/0003-4916(90)90368-X
|
| [62] |
Vitiello G (2000) Defect formation through boson condensation in quantum field theory. Topological Defects and the Non-Equilibrium Dynamics of Symmetry Breaking Phase Transitions Dordrecht: Springer, 171-191. doi: 10.1007/978-94-011-4106-2_9
|
| [63] |
Meyer G, Vitiello G (2018) On the molecular dynamics in the hurricane interactions with its environment. Phys Lett A 382: 1441-1448. doi: 10.1016/j.physleta.2018.03.044
|
| [64] |
Da Silva AF, Carpenter T, How TV, et al. (1997) Stable vortices within vein cuffs inhibit anastomotic myointimal hyperplasia? Eur J Vasc Endovasc Surg 14: 157-163. doi: 10.1016/S1078-5884(97)80185-2
|
| [65] |
Kefayati S, Amans M, Faraji F, et al. (2017) The manifestation of vortical and secondary flow in the cerebral venous outflow tract: An in vivo MR velocimetry study. J Biomech 50: 80-187. doi: 10.1016/j.jbiomech.2016.11.041
|
| [66] |
Lurie F, Kistner RL, Eklof B, et al. (2003) Mechanism of venous valve closure and role of the valve in circulation: a new concept. J Vasc Surg 38: 955-961. doi: 10.1016/S0741-5214(03)00711-0
|
| [67] | Boisseau MR (1997) Venous valves in the legs: hemodynamic and biological problems and relationship to physiopathology. J Mal Vasc 22: 122-127. |
| [68] |
Machi J, Sigel B, Ramos JR, et al. (1986) Sonographic evaluation of platelet aggregate retention in a vortex within a simulated venous sinus. J Ultrasound Med 5: 685-689. doi: 10.7863/jum.1986.5.12.685
|
| [69] |
Meyer G, Vitiello G (2019) On the hurricane collective molecular dynamics. J Phys Conf Ser 1275: 012017. doi: 10.1088/1742-6596/1275/1/012017
|
| [70] |
Higgs PW (1966) Spontaneous symmetry breakdown without massless bosons. Phys Rev 145: 1156-1163. doi: 10.1103/PhysRev.145.1156
|
| [71] |
Freeman W, Vitiello G (2010) Vortices in brain waves. Int J Mod Phys B 24: 3269-3295. doi: 10.1142/S0217979210056025
|
| [72] | Del Giudice E, Vitiello G (2006) The role of the electromagnetic field in the formation of domains in the process of symmetry breaking phase transitions. Phys Rev A 74: 02210. |
| [73] | De Lauro E, De Martino S (2019) On the heart vibrations: Some insights from ECG and laser doppler vibrometry.p. atticon11705 V-C-5in press. |
| [74] | Scalise L (2012) Non contact heart monitoring. Advances in Electrocardiograms-Methods and Analysis Rijeka, Croatia: InTech, 81-106. |
| [75] | Tomasini E, Pinotti M, Paone N (1998) Carotid artery pulse wave measured by a laser vibrometer.3411: 611-616. |
| [76] |
Morbiducci U, Scalise L, De Melis M (2006) Optical vibrocardiography: a novel tool for optical monitoring of cardiac activity. Ann Biomed Eng 35: 45-58. doi: 10.1007/s10439-006-9202-9
|
| [77] |
Buccheri G, De Lauro E, De Martino S, et al. (2016) Experimental study of self-oscillations of the trachea–larynx tract by laser doppler vibrometry. Biomed Phys Eng Express 2: 055009. doi: 10.1088/2057-1976/2/5/055009
|
| [78] | Tavakolian K, Dumont GA, Blaber AP (2012) Analysis of seismocardiogram capability for trending stroke volume changes: A lower body negative pressure study. Computing in Cardiology 2012: 733-736. |
| [79] | Desjardins CL, Antonelli LT (2007) A remote and non-contact method for obtaining the blood pulse waveform with a laser Doppler vibrometer. Advanced Biomedical and Clinical Diagnostic Systems V, Proc. SPIE . |
| [80] |
Hyvärinen A, Karhunen J, Oja E (2001) Independent Component Analysis Hoboken, USA: John Wiley & Sons. doi: 10.1002/0471221317
|
| [81] |
Capuano P, De Lauro E, De Martino S, et al. (2017) Convolutive independent component analysis for processing massive datasets: a case study at Campi Flegrei (Italy). Nat Hazards 86: 417-429. doi: 10.1007/s11069-016-2545-0
|
| [82] |
Capuano P, De Lauro E, De Martino S (2016) Detailed investigation of long-period activity at Campi Flegrei by convolutive independent component analysis. Phys Earth Planet Inter 253: 48-57. doi: 10.1016/j.pepi.2016.02.003
|
| [83] |
Capuano P, De Lauro E, De Martino S (2011) Water-level oscillations in the Adriatic Sea as coherent self-oscillations inferred by independent component analysis. Prog Oceanogr 91: 447-460. doi: 10.1016/j.pocean.2011.06.001
|
| [84] |
De Lauro E, De Martino S, Falanga M, et al. (2006) Statistical analysis of Stromboli VLP tremor in the band [0.1–0.5] Hz: some consequences for vibrating structures. Nonlinear Process Geophys 13: 393-400. doi: 10.5194/npg-13-393-2006
|
| [85] |
Rüssel IK, Götte MJW, Bronzwaer JG, et al. (2009) Left ventricular torsion: an expanding role in the analysis of myocardial dysfunction. JACC Cardiovasc Imaging 2: 648-655. doi: 10.1016/j.jcmg.2009.03.001
|
| [86] |
Loppini A, Capolupo A, Cherubini C (2012) On the coherent behavior of pancreatic beta cell clusters. Phys Lett A 378: 3210-3217. doi: 10.1016/j.physleta.2014.09.041
|
| [87] |
Dal Lin C, Brugnolo L, Marinova M, et al. (2020) Toward a unified view of cognitive and biochemical activity: Meditation and linguistic self-reconstructing may lead to inflammation and oxidative stress improvement. Entropy 22: 818. doi: 10.3390/e22080818
|
| [88] | Dal Lin C, Radu CM, Vitiello G, et al. (2020) In vitro effects on cellular shaping, contratility, cytoskeletal organization and mitochondrial activity in HL1 cells after different sounds stimulation. A qualitative pilot study and a theoretical physical model, 2020. |
| [89] | McFadden J, Al-Khalili J (2014) Life on the edge: the coming of age of quantum biology London: Bantam Press. |
| [90] |
McFadden J, Al-Khalili J (1999) A quantum mechanical model of adaptive mutation. Biosystems 50: 203-211. doi: 10.1016/S0303-2647(99)00004-0
|
| [91] |
Misra B, Sudarshan ECG (1977) The Zeno's paradox in quantum theory. J Math Phys 18: 756-763. doi: 10.1063/1.523304
|
| [92] |
Kraus K (1981) Measuring processes in quantum mechanics I. Continuous observation and the watchdog effect. Found Phys 11: 547-576. doi: 10.1007/BF00726936
|
| [93] |
Meloni M (2014) The social brain meets the reactive genome: neuroscience, epigenetics and the new social biology. Front Hum Neurosci 8: 309. doi: 10.3389/fnhum.2014.00309
|
| [94] |
Vitiello G (2014) On the isomorphism between dissipative systems, fractal self-similarity and electrodynamics. Toward an integrated vision of nature. Systems 2: 203-216. doi: 10.3390/systems2020203
|
| [95] |
Novack DH, Cameron O, Epel E, et al. (2007) Psychosomatic medicine: The scientific foundation of the biopsychosocial model. Acad Psychiatry 31: 388-401. doi: 10.1176/appi.ap.31.5.388
|
| [96] |
Dal Lin C, Gola E, Brocca A, et al. (2018) miRNAs may change rapidly with thoughts: The relaxation response after myocardial infarction. Eur J Integr Med 20: 63-72. doi: 10.1016/j.eujim.2018.03.009
|
| [97] |
Chrousos GP (2009) Stress and disorders of the stress system. Nat Rev Endocrinol 5: 374-381. doi: 10.1038/nrendo.2009.106
|
| [98] |
Muehsam D, Ventura C (2014) Life rhythm as a symphony of oscillatory patterns: electromagnetic energy and sound vibration modulates gene expression for biological signaling and healing. Glob Adv Health Med 3: 40-55. doi: 10.7453/gahmj.2014.008
|
| [99] | (2015) Heart Beat Made Visible on CymaScope Avaibale from: https://www.youtube.com/watch?v=2kuY98F7o_0. |
| [100] |
Ingber DE, Wang N, Stamenović D (2014) Tensegrity, cellular biophysics, and the mechanics of living systems. Rep Prog Phys 77: 046603. doi: 10.1088/0034-4885/77/4/046603
|
| [101] |
Wang N, Tytell JD, Ingber DE (2009) Mechanotransduction at a distance: Mechanically coupling the extracellular matrix with the nucleus. Nat Rev Mol Cell Biol 10: 75-82. doi: 10.1038/nrm2594
|
| [102] |
Martino F, Perestrelo AR, Vinarsky V, et al. (2018) Cellular mechanotransduction: from tension to function. Frony Physiol 9: 824. doi: 10.3389/fphys.2018.00824
|
| [103] |
Buxbaum O (2016) Key Insights into Basic Mechanisms of Mental Activity Switzerland: Springer International Publishing. doi: 10.1007/978-3-319-29467-4
|
| [104] |
Jamieson JP, Crum AJ, Goyer JP, et al. (2018) Optimizing stress responses with reappraisal and mindset interventions: an integrated model. Anxiety, Stress, Coping 31: 245-261. doi: 10.1080/10615806.2018.1442615
|
| [105] |
Pulvermüller F (2013) How neurons make meaning: Brain mechanisms for embodied and abstract-symbolic semantics. Trends Cogn Sci 17: 458-470. doi: 10.1016/j.tics.2013.06.004
|
| [106] |
Segall JM, Allen EA, Jung RE, et al. (2012) Correspondence between structure and function in the human brain at rest. Front Neuroinform 6: 10. doi: 10.3389/fninf.2012.00010
|
| [107] |
Alexander-Bloch A, Shou H, Liu S, et al. (2018) On testing for spatial correspondence between maps of human brain structure and function. Neuroimage 178: 540-551. doi: 10.1016/j.neuroimage.2018.05.070
|
| [108] |
Rebollo I, Devauchelle AD, Béranger B, et al. (2018) Stomach-brain synchrony reveals a novel, delayed-connectivity resting-state network in humans. Elife 7: e33321. doi: 10.7554/eLife.33321
|
| [109] | Ventura C (2017) Seeing cell biology with the eyes of physics. NanoWorld J 3: S1-S8. |
| [110] | Fredericks S, Saylor JR (2013) Shape oscillation of a levitated drop in an acoustic field, 2013 Available from: arXiv:1310.2967. |
| [111] |
Zhang CY, Wang Y, Schubert R, et al. (2016) Effect of audible sound on protein crystallization. Cryst Growth Des 16: 705-713. doi: 10.1021/acs.cgd.5b01268
|
| [112] |
Guo F, Li P, French JB, et al. (2015) Controlling cell–cell interactions using surface acoustic waves. Proc Natl Acad Sci 112: 43-48. doi: 10.1073/pnas.1422068112
|
| [113] |
Vogel V, Sheetz M (2006) Local force and geometry sensing regulate cell functions. Nat Rev Mol Cell Biol 7: 265-275. doi: 10.1038/nrm1890
|
| [114] |
Shaobin G, Wu Y, Li K, et al. (2010) A pilot study of the effect of audible sound on the growth of Escherichia coli. Colloid Surface B 78: 367-371. doi: 10.1016/j.colsurfb.2010.02.028
|
| [115] |
Gu SB, Yang B, Wu Y, et al. (2013) Growth and physiological characteristics of E. coli in response to the exposure of sound field. Pakistan J Biol Sci 16: 969-975. doi: 10.3923/pjbs.2013.969.975
|
| [116] |
Sahu S, Ghosh S, Fujita D, et al. (2014) Live visualizations of single isolated tubulin protein self-assembly via tunneling current: effect of electromagnetic pumping during spontaneous growth of microtubule. Sci Rep 4: 7303. doi: 10.1038/srep07303
|
| [117] |
Acbas G, Niessen KA, Snell EH, et al. (2014) Optical measurements of long-range protein vibrations. Nat Commun 5: 3076. doi: 10.1038/ncomms4076
|
| [118] |
Christians ES, Benjamin IJ (2012) Proteostasis and REDOX state in the heart. Am J Physiol Heart Circ Physiol 302: H24-H37. doi: 10.1152/ajpheart.00903.2011
|
| [119] |
Christians ES, Mustafi SB, Benjamin IJ (2014) Chaperones and cardiac misfolding protein diseases. Curr Protein Pept Sci 15: 189-204. doi: 10.2174/1389203715666140331111518
|
| [120] |
Naviaux RK (2014) Metabolic features of the cell danger response. Mitochondrion 16: 7-17. doi: 10.1016/j.mito.2013.08.006
|
| [121] |
Crum A, Zuckerman B (2017) Changing mindsets to enhance treatment effectiveness. J Am Med Assoc 317: 2063-2064. doi: 10.1001/jama.2017.4545
|
| [122] |
Maas C, Belgardt D, Han KL, et al. (2009) Synaptic activation modifies microtubules underlying transport of postsynaptic cargo. Proc Natl Acad Sci 106: 8731-8736. doi: 10.1073/pnas.0812391106
|
| [123] |
Lo LP, Liu SH, Chang YC (2007) Assembling microtubules disintegrate the postsynaptic density in vitro. Cell Motil Cytoskeleton 64: 6-18. doi: 10.1002/cm.20163
|
| [124] |
Arimura N, Kaibuchi K (2007) Neuronal polarity: From extracellular signals to intracellular mechanisms. Nat Rev Neurosci 8: 194-205. doi: 10.1038/nrn2056
|
| [125] |
Macario AJL, Conway de Macario E (2000) Stress and molecular chaperones in disease. Int J Clin Lab Res 30: 49-66. doi: 10.1007/s005990070016
|
| [126] |
Dal Lin C, Marinova M, Brugnolo L, et al. Rapid senectome and alternative splicing miRNAs changes with the relaxation response: A one year follow-up study, 2020 Available from: |
| [127] |
Picard M, McManus MJ, Gray JD, et al. (2015) Mitochondrial functions modulate neuroendocrine, metabolic, inflammatory, and transcriptional responses to acute psychological stress. Proc Natl Acad Sci 112: E6614-E6623. doi: 10.1073/pnas.1515733112
|
| [128] | Piattelli-Palmarini M, Vitiello G (2015) Linguistics and some aspects of its underlying dynamics. Biolinguistics 9: 96-115. |
| [129] |
Mańka R, Ogrodnik B (1991) A model of soliton transport along microtubules. J Biol Phys 18: 85-189. doi: 10.1007/BF00417807
|
| [130] |
Kučera O, Havelka D (2012) Mechano-electrical vibrations of microtubules-Link to subcellular morphology. BioSystems 109: 346-355. doi: 10.1016/j.biosystems.2012.04.009
|
| [131] |
Benias PC, Wells RG, Sackey-Aboagye B, et al. (2018) Structure and distribution of an unrecognized interstitium in human tissues. Sci Rep 8: 4947. doi: 10.1038/s41598-018-23062-6
|
| [132] |
Brizhik L, Chiappini E, Stefanini P, et al. (2019) Modeling meridians within the quantum field theory. J Acupunct Meridian Stud 12: 29-36. doi: 10.1016/j.jams.2018.06.009
|
| [133] | Bentov I (1977) Stalking the wild pendulum Glasgow: William Collins Sons & Co. Ltd. |
| [134] |
Pavanello S, Campisi m, Tona F, et al. (2019) Exploring epigenetic age in response to intensive relaxing training: A pilot study to slow down biological age. Int J Env Res Pub He 16: 3074. doi: 10.3390/ijerph16173074
|
| [135] | Dal Lin C, Grasso R, Scordino A, et al. (2020) Ph, electric conductivity and delayed luminescence changes in human sera of subjects undergoing the relaxation response: A pilot study Available from: doi:10.20944/PREPRINTS202004.0202.V1. |
| [136] |
Cifra M, Brouder C, Nerudová M, et al. (2015) Biophotons, coherence and photocount statistics: A critical review. J Lumin 164: 38-51. doi: 10.1016/j.jlumin.2015.03.020
|
| [137] |
Boveris A, Cadenas E, Reiter R (1980) Organ chemiluminescence: noninvasive assay for oxidative radical reactions. Proc Natl Acad Sci 77: 347-351. doi: 10.1073/pnas.77.1.347
|
| [138] |
Krasovitski B, Frenkel V, Shoham S (2011) Intramembrane cavitation as a unifying mechanism for ultrasound-induced bioeffects. Proc Natl Acad Sci 108: 3258-3263. doi: 10.1073/pnas.1015771108
|
| [139] |
Brujan EA (2000) Collapse of cavitation bubbles in blood. Europhys Lett 50: 175. doi: 10.1209/epl/i2000-00251-7
|
| [140] |
Brennen CE (2015) Cavitation in medicine. Interface Focus 5: 20150022. doi: 10.1098/rsfs.2015.0022
|
| [141] |
Didenko YT, Suslick KS (2002) The energy efficiency of formation of photons radicals and ions during single-bubble cavitation. Nature 418: 394-397. doi: 10.1038/nature00895
|
| [142] |
Sabbadini SA, Vitiello G (2019) Entanglement and phase-mediated correlations in quantum field theory. Application to brain-mind states. Appl Sci 9: 3203. doi: 10.3390/app9153203
|
| [143] |
Shaffer F, McCraty R, Zerr CL (2014) A healthy heart is not a metronome: an integrative review of the heart's anatomy and heart rate variability. Front Psychol 5: 1040. doi: 10.3389/fpsyg.2014.01040
|
| [144] |
Grippo A (2011) The utility of animal models in understanding links between psychosocial processes and cardiovascular health. Soc Pers Psychol Compass 5: 164-179. doi: 10.1111/j.1751-9004.2011.00342.x
|
| [145] |
Mensah G, Collins P (2015) Understanding mental health for the prevention and control of cardiovascular diseases. Glob Heart 10: 221. doi: 10.1016/j.gheart.2015.08.003
|
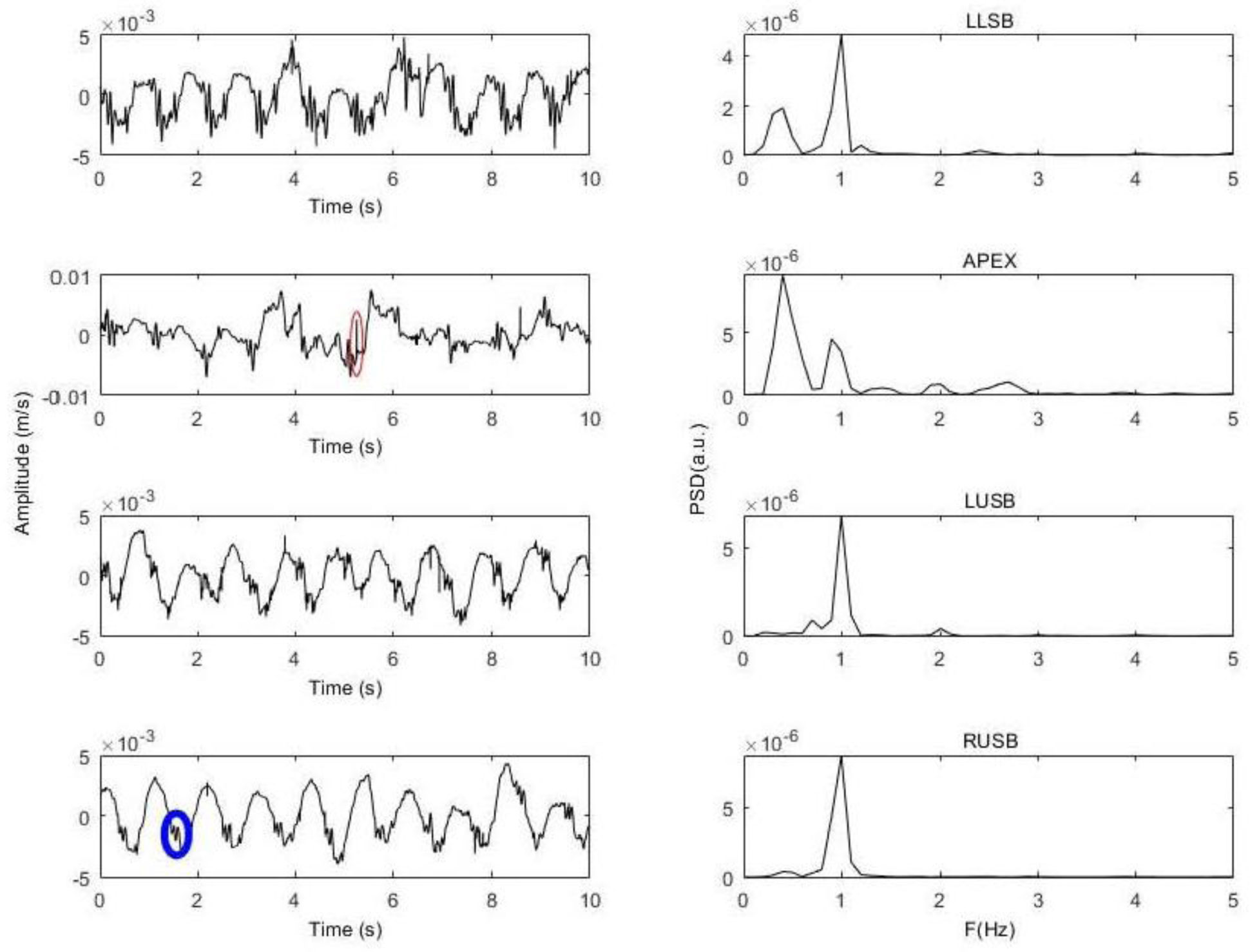

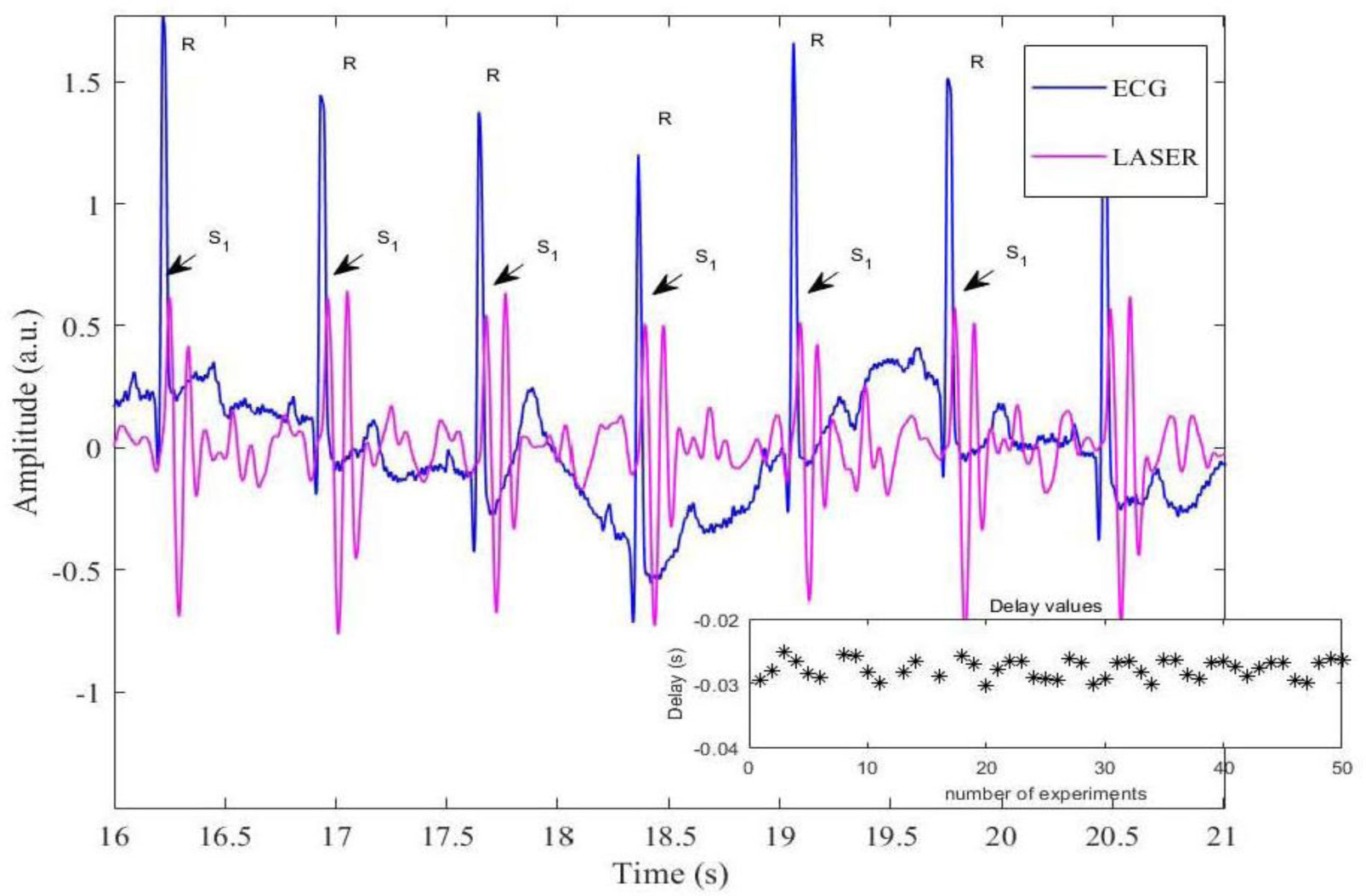
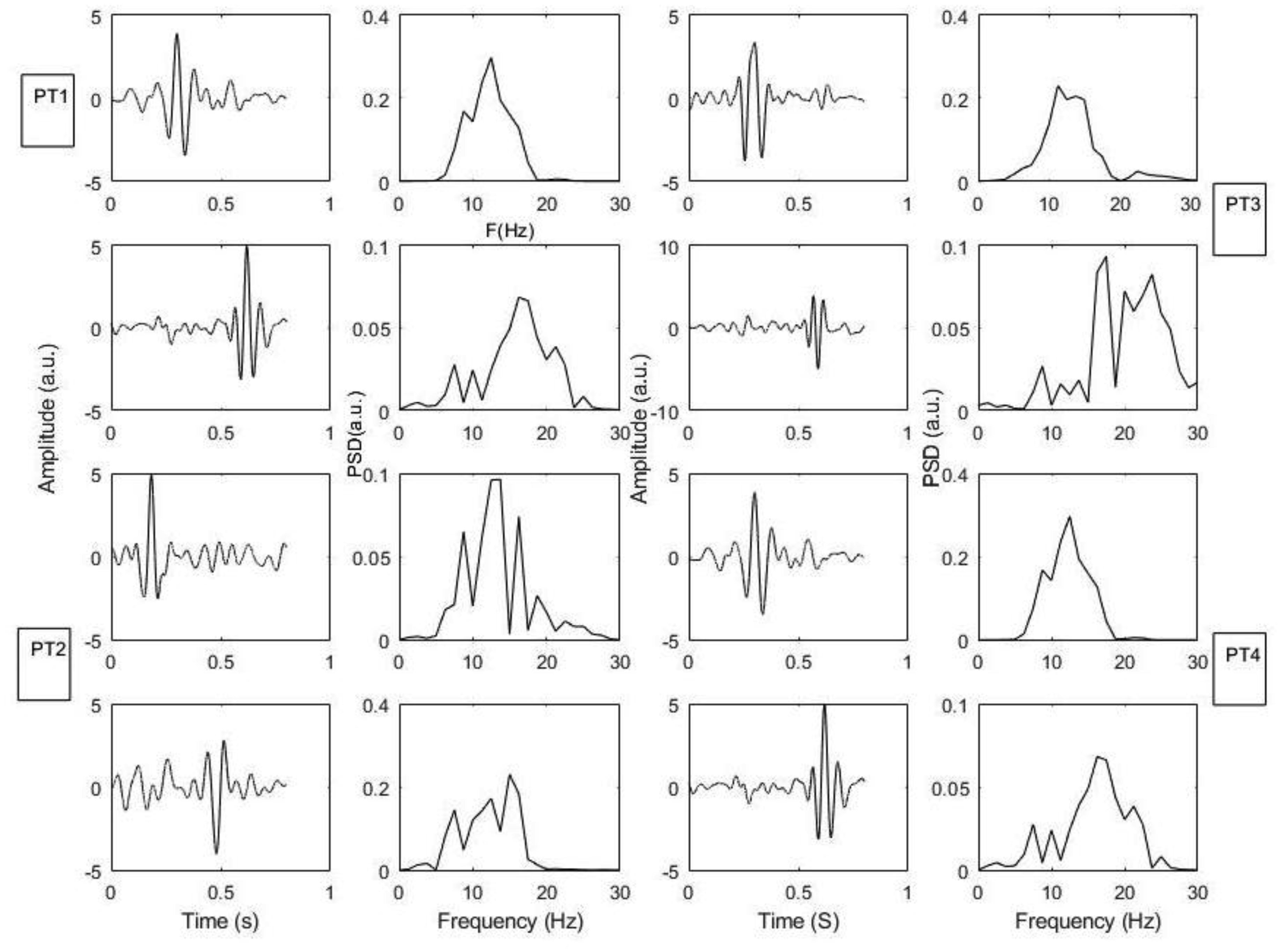
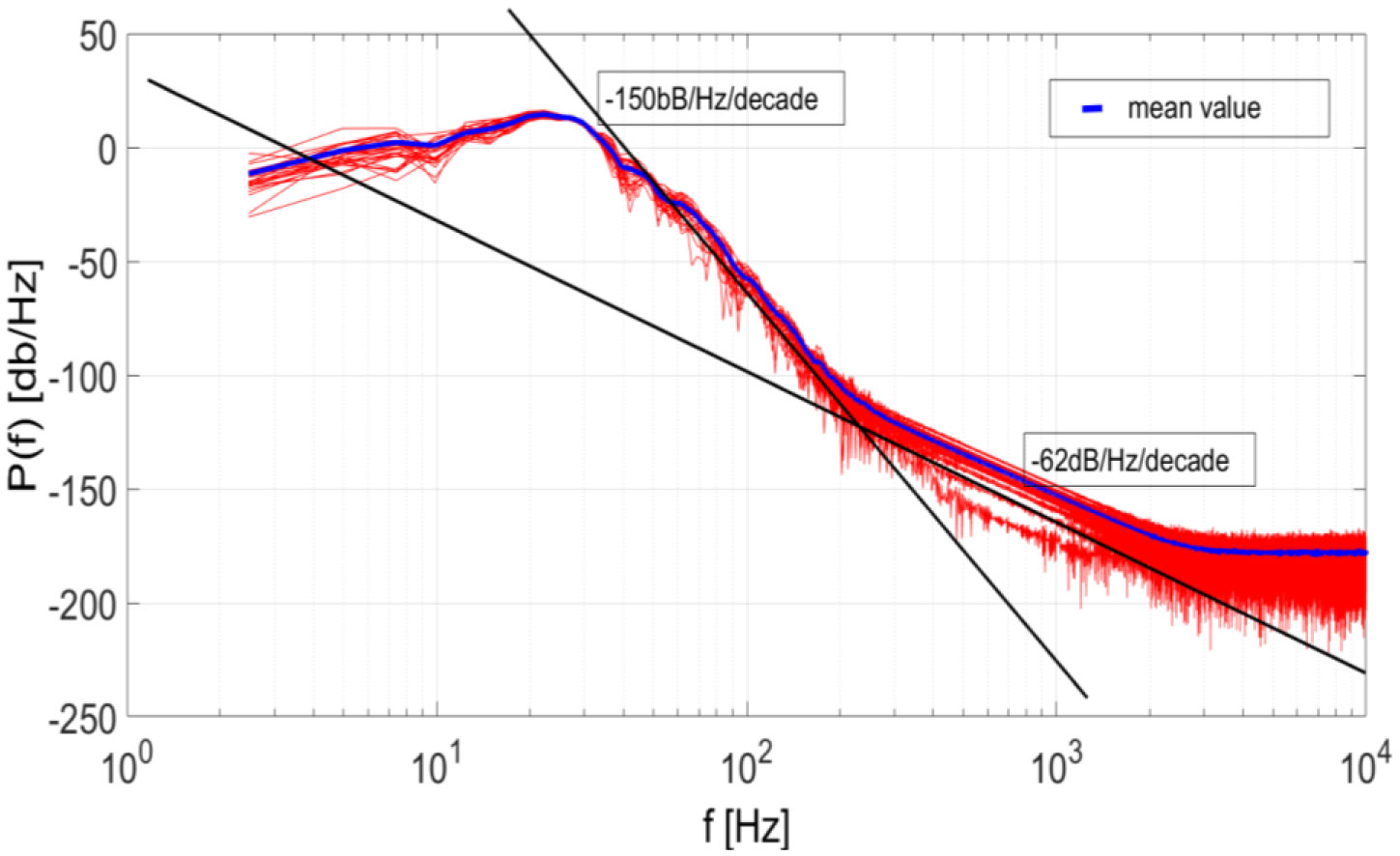
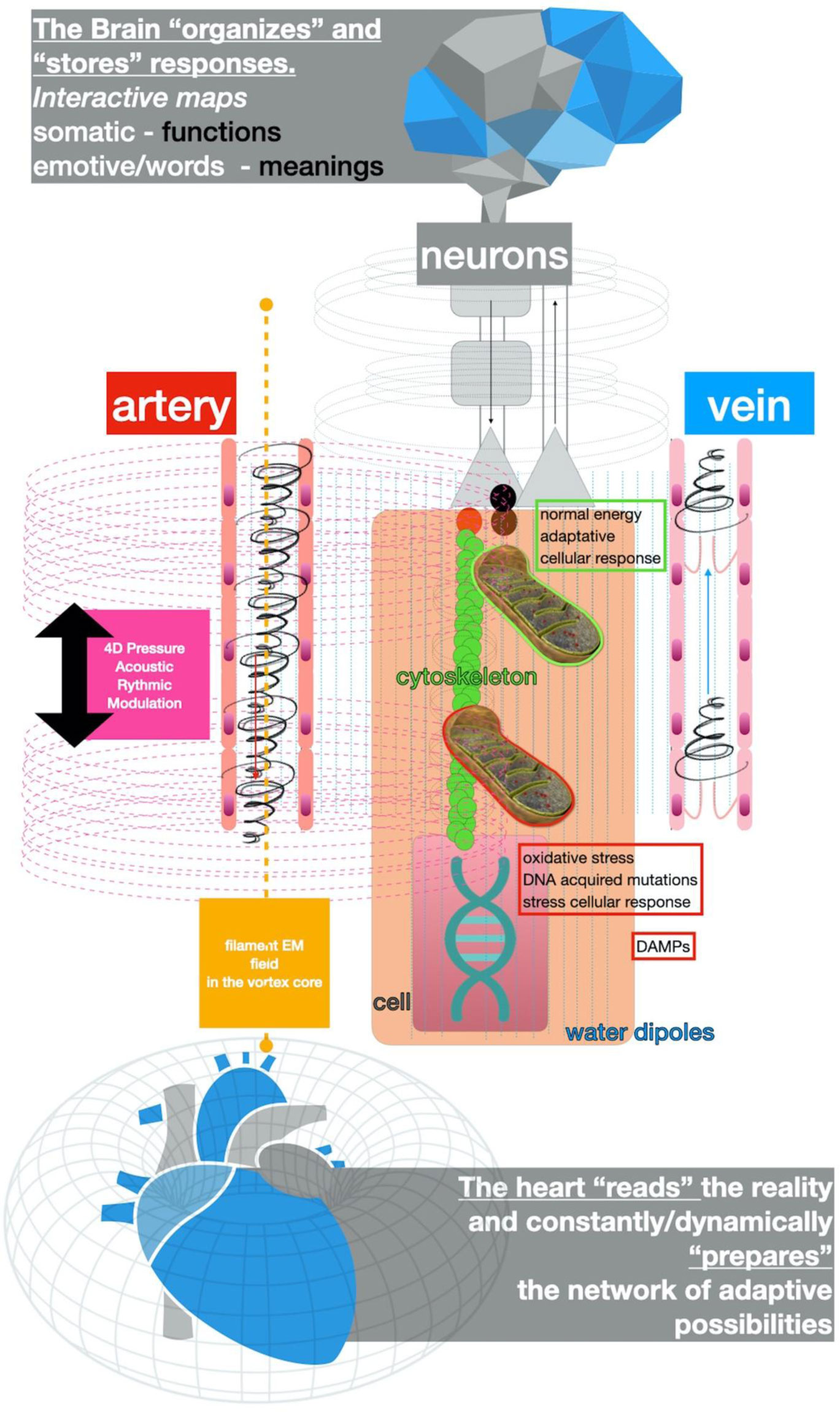

Figures(7)

C. Dal Lin, M. Falanga, E. De Lauro, S. De Martino, G. Vitiello. Biochemical and biophysical mechanisms underlying the heart and the brain dialog[J]. AIMS Biophysics, 2021, 8(1): 1-33. doi: 10.3934/biophy.2021001







 DownLoad:
DownLoad: