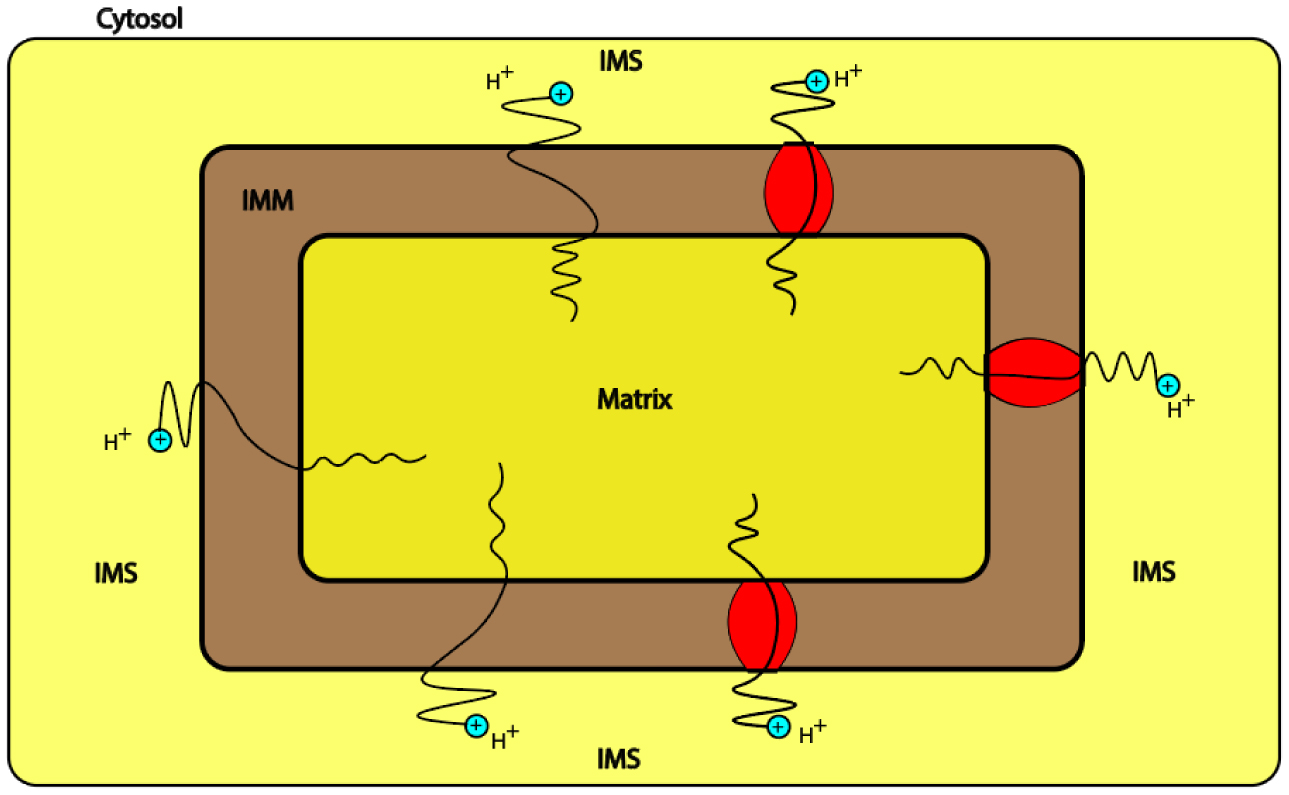
The mitochondrion is a vital intracellular organelle that is responsible for ATP production. It utilizes both the concentration gradient and the electrical potential of the inner mitochondrial membrane to drive the flow of protons from the intermembrane space to the matrix to generate ATP via ATP-synthase. However, the proton leak flow, which is mediated via the inner mitochondrial membrane and uncoupling proteins, can reduce the efficiency of ATP production. Protons can exhibit a quantum behavior within biological systems. However, the investigation of the quantum behavior of protons within the mitochondria is lacking particularly in the contribution to the proton leak. In the present study, we proposed a mathematical model of protons tunneling through the inner mitochondrial membrane and the mitochondrial carrier superfamily MCF including uncoupling proteins UCPs and the adenine nucleotide translocases ANTs. According to the model and its assumptions, the quantum tunneling of protons may contribute significantly to the proton leak if it is compared with the classical flow of protons. The quantum tunneling proton leak may depolarize the membrane potential, hence it may contribute to the physiological regulation of ATP synthesis and reactive oxygen species ROS production. In addition to that, the mathematical model of proton tunneling suggested that the proton-tunneling leak may depolarize the membrane potential to values beyond the physiological needs which in turn can harm the mitochondria and the cells. Moreover, we argued that the quantum proton leak might be more energetically favorable if it is compared with the classical proton leak. This may give the advantage for quantum tunneling of protons to occur since less energy is required to contribute significantly to the proton leak compared with the classical proton flow.
Citation: Mahmoud Abdallat, Abdallah Barjas Qaswal, Majed Eftaiha, Abdel Rahman Qamar, Qusai Alnajjar, Rawand Sallam, Lara Kollab, Mohammad Masa'deh, Anas Amayreh, Hiba Mihyar, Hesham Aboushakra, Bayan Alkelani, Rawan Owaimer, Mohannad Abd-Alhadi, Salwa Ireiqat, Fahed Turk, Ahmad Daoud, Bashar Darawsheh, Ahmad Hiasat, Majd Alhalaki, Shahem Abdallat, Salsabiela Bani Hamad, Rand Murshidi. A mathematical modeling of the mitochondrial proton leak via quantum tunneling[J]. AIMS Biophysics, 2024, 11(2): 189-233. doi: 10.3934/biophy.2024012
The mitochondrion is a vital intracellular organelle that is responsible for ATP production. It utilizes both the concentration gradient and the electrical potential of the inner mitochondrial membrane to drive the flow of protons from the intermembrane space to the matrix to generate ATP via ATP-synthase. However, the proton leak flow, which is mediated via the inner mitochondrial membrane and uncoupling proteins, can reduce the efficiency of ATP production. Protons can exhibit a quantum behavior within biological systems. However, the investigation of the quantum behavior of protons within the mitochondria is lacking particularly in the contribution to the proton leak. In the present study, we proposed a mathematical model of protons tunneling through the inner mitochondrial membrane and the mitochondrial carrier superfamily MCF including uncoupling proteins UCPs and the adenine nucleotide translocases ANTs. According to the model and its assumptions, the quantum tunneling of protons may contribute significantly to the proton leak if it is compared with the classical flow of protons. The quantum tunneling proton leak may depolarize the membrane potential, hence it may contribute to the physiological regulation of ATP synthesis and reactive oxygen species ROS production. In addition to that, the mathematical model of proton tunneling suggested that the proton-tunneling leak may depolarize the membrane potential to values beyond the physiological needs which in turn can harm the mitochondria and the cells. Moreover, we argued that the quantum proton leak might be more energetically favorable if it is compared with the classical proton leak. This may give the advantage for quantum tunneling of protons to occur since less energy is required to contribute significantly to the proton leak compared with the classical proton flow.
ATP
pmf
ANT
UCP
ROS
ADT
GTP
GDP
MCF
IMM
PMF
IMS
ETC

| [1] |
Picard M, Taivassalo T, Gouspillou G, et al. (2011) Mitochondria: isolation, structure and function. J Physiol 589: 4413-4421. https://doi.org/10.1113/jphysiol.2011.212712 
|
| [2] |
Duchen MR (2004) Roles of mitochondria in health and disease. Diabetes 1: S96-102. https://doi.org/10.2337/diabetes.53.2007.S96
|
| [3] |
Ardalan A, Smith MD, Jelokhani-Niaraki M (2022) Uncoupling proteins and regulated proton leak in mitochondria. Int J Mol Sci 23: 1528. https://doi.org/10.3390/ijms23031528
|
| [4] |
Nesci S (2023) Proton leak through the UCPs and ANT carriers and beyond: a breath for the electron transport chain. Biochimie 214: 77-85. https://doi.org/10.1016/j.biochi.2023.06.008
|
| [5] |
Jastroch M, Divakaruni AS, Mookerjee S, et al. (2010) Mitochondrial proton and electron leaks. Essays Biochem 47: 53-67. https://doi.org/10.1042/bse0470053
|
| [6] |
Bertholet AM, Kirichok Y (2022) Mitochondrial H+ leak and thermogenesis. Annu Rev Physiol 84: 381-407. https://doi.org/10.1146/annurev-physiol-021119-034405
|
| [7] |
Nicholls DG (1997) The non-Ohmic proton leak—25 years on. Biosci Rep 17: 251-257. https://doi.org/10.1023/A:1027376426860
|
| [8] |
Rupprecht A, Sokolenko EA, Beck V, et al. (2010) Role of the transmembrane potential in the membrane proton leak. Biophys J 98: 1503-1511. https://doi.org/10.1016/j.bpj.2009.12.4301
|
| [9] |
Kenwood BM, Weaver JL, Bajwa A, et al. (2014) Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Mol Metab 3: 114-123. https://doi.org/10.1016/j.molmet.2013.11.005
|
| [10] |
Sack MN (2006) Mitochondrial depolarization and the role of uncoupling proteins in ischemia tolerance. Cardiovasc Res 72: 210-219. https://doi.org/10.1016/j.cardiores.2006.07.010
|
| [11] |
Padmaraj D, Pande R, Miller JH, et al. (2014) Mitochondrial membrane studies using impedance spectroscopy with parallel pH monitoring. PLoS One 9: e101793. https://doi.org/10.1371/journal.pone.0101793
|
| [12] | Serway RA, Kirkpatrick LD Physics for Scientists and Engineers with Modern Physics James Madison University (2005). https://doi.org/10.1119/1.2342517 |
| [13] |
Calvillo L, Redaelli V, Ludwig N, et al. (2022) Quantum biology research meets pathophysiology and therapeutic mechanisms: a biomedical perspective. Quantum Rep 4: 148-712. https://doi.org/10.3390/quantum4020011
|
| [14] |
Kim Y, Bertagna F, D'souza EM, et al. (2021) Quantum biology: an update and perspective. Quantum Rep 3: 80-126. https://doi.org/10.3390/quantum3010006
|
| [15] |
Bennett JP, Onyango IG (2021) Energy, entropy and quantum tunneling of protons and electrons in brain mitochondria: relation to mitochondrial impairment in aging-related human brain diseases and therapeutic measures. Biomedicines 9: 225. https://doi.org/10.3390/biomedicines9020225
|
| [16] |
Nunn AV, Guy GW, Bell JD (2022) Thermodynamics and inflammation: insights into quantum biology and ageing. Quantum Rep 4: 47-74. https://doi.org/10.3390/quantum4010005
|
| [17] |
Slocombe L, Sacchi M, Al-Khalili J (2022) An open quantum systems approach to proton tunnelling in DNA. Commun Phys 5: 109. https://doi.org/10.1038/s42005-022-00881-8
|
| [18] |
Çelebi G, Özçelik E, Vardar E, et al. (2021) Time delay during the proton tunneling in the base pairs of the DNA double helix. Prog Biophys Mol Biol 167: 96-103. https://doi.org/10.1016/j.pbiomolbio.2021.06.001
|
| [19] |
Layfield JP, Hammes-Schiffer S (2014) Hydrogen tunneling in enzymes and biomimetic models. Chem Rev 114: 3466-3494. https://doi.org/10.1021/cr400400p
|
| [20] |
Xin H, Sim WJ, Namgung B, et al. (2019) Quantum biological tunnel junction for electron transfer imaging in live cells. Nat Commun 10: 3245. https://doi.org/10.1038/s41467-019-11212-x
|
| [21] |
Hayashi T, Stuchebrukhov AA (2010) Electron tunneling in respiratory complex I. Proc Natl Acad Sci USA 107: 19157-19162. https://doi.org/10.1073/pnas.1009181107
|
| [22] |
Hagras MA, Hayashi T, Stuchebrukhov AA (2015) Quantum calculations of electron tunneling in respiratory complex III. J Phys Chem B 119: 14637-14651. https://doi.org/10.1021/acs.jpcb.5b09424
|
| [23] |
Gray HB, Winkler JR (2003) Electron tunneling through proteins. Q Rev Biophys 36: 341-372. https://doi.org/10.1017/S0033583503003913
|
| [24] |
Qaswal AB (2019) Quantum tunneling of ions through the closed voltage-gated channels of the biological membrane: A mathematical model and implications. Quantum Rep 1: 219-225. https://doi.org/10.3390/quantum1020019
|
| [25] |
Qaswal AB, Ababneh O, Khreesha L, et al. (2021) Mathematical modeling of ion quantum tunneling reveals novel properties of voltage-gated channels and quantum aspects of their pathophysiology in excitability-related disorders. Pathophysiology 28: 116-154. https://doi.org/10.3390/pathophysiology28010010
|
| [26] |
Vorobyov I, Olson TE, Kim JH, et al. (2014) Ion-induced defect permeation of lipid membranes. Biophys. J 106: 586-597. https://doi.org/10.1016/j.bpj.2013.12.027
|
| [27] |
Aryal P, Sansom MS, Tucker SJ (2015) Hydrophobic gating in ion channels. J Mol Biol 427: 121-130. https://doi.org/10.1016/j.jmb.2014.07.030
|
| [28] |
Khavrutskii IV, Gorfe AA, Lu B (2009) Free energy for the permeation of Na+ and Cl− ions and their ion-pair through a zwitterionicdimyristoyl phosphatidylcholine lipid bilayer by umbrella integration with harmonic fourier beads. J Am Chem Soc 131: 1706-1716. https://doi.org/10.1021/ja8081704
|
| [29] |
Klesse G, Tucker SJ, Sansom MS (2020) Electric field induced wetting of a hydrophobic gate in a model nanopore based on the 5-HT3 receptor channel. ACS Nano 14: 10480-10491. https://doi.org/10.1021/acsnano.0c04387
|
| [30] |
Miyazaki T (2004) Atom Tunneling Phenomena in Physics, Chemistry and Biology. Berlin: Springer Science & Business Media. https://doi.org/10.1007/978-3-662-05900-5
|
| [31] | Bertil H, Bertil H (2001) Ion Channels of Excitable Membranes. Sunderland: Sinauer Associates. https://doi.org/10.1016/0014-5793(92)81020-m |
| [32] |
Lee JW (2019) Electrostatically localized proton bioenergetics: better understanding membrane potential. Heliyon 5: e01961. https://doi.org/10.1016/j.heliyon.2019.e01961
|
| [33] |
Silverstein TP (2022) A critique of the capacitor-based “transmembrane electrostatically localized proton” hypothesis. J Bioenerg Biomembr 54: 59-65. https://doi.org/10.1007/s10863-022-09931-w
|
| [34] |
Qaswal AB (2020) Quantum electrochemical equilibrium: quantum version of the Goldman–Hodgkin–Katz equation. Quantum Rep 2: 266-277. https://doi.org/10.3390/quantum2020017
|
| [35] |
Lee JW (2020) Protonic capacitor: elucidating the biological significance of mitochondrial cristae formation. Sci Rep 10: 10304. https://doi.org/10.1038/s41598-020-66203-6
|
| [36] |
Lee C, Wallace DC, Burke PJ (2023) Super-resolution imaging of voltages in the interior of individual, vital mitochondria. Acs Nano 18: 1345-1356. https://doi.org/10.1021/acsnano.3c02768
|
| [37] |
Nicholls D (2002) Mitochondrial bioenergetics, aging, and aging-related disease. Sci Aging Knowl Environ 2002: pe12. https://doi.org/10.1126/sageke.2002.31.pe12
|
| [38] |
Norat P, Soldozy S, Sokolowski JD, et al. (2020) Mitochondrial dysfunction in neurological disorders: Exploring mitochondrial transplantation. NPJ Regen Med 5: 22. https://doi.org/10.1038/s41536-020-00107-x
|
| [39] |
Brookes PS, Land JM, Clark JB, et al. (1998) Peroxynitrite and brain mitochondria: evidence for increased proton leak. J Neurochem 70: 2195-2202. https://doi.org/10.1046/j.1471-4159.1998.70052195.x
|
| [40] |
Xie P, Zhang H, Qin Y, et al. (2023) Membrane proteins and membrane curvature: mutual interactions and a perspective on disease treatments. Biomolecules 13: 1772. https://doi.org/10.3390/biom13121772
|
| [41] |
Morris CE (2011) Voltage-gated channel mechanosensitivity: fact or friction?. Front Physiol 2: 25. https://doi.org/10.3389/fphys.2011.00025
|
| [42] |
Cortassa S, O'Rourke B, Aon MA (2014) Redox-optimized ROS balance and the relationship between mitochondrial respiration and ROS. Biochim Biophys Acta 1837: 287-295. https://doi.org/10.1016/j.bbabio.2013.11.007
|
| [43] |
vanHameren G, Campbell G, Deck M, et al. (2019) In vivo real-time dynamics of ATP and ROS production in axonal mitochondria show decoupling in mouse models of peripheral neuropathies. Acta Neuropathol Commun 7: 86. https://doi.org/10.1186/s40478-019-0740-4
|
| [44] |
Berry BJ, Kaeberlein M (2021) An energetics perspective on geroscience: mitochondrial protonmotive force and aging. Geroscience 43: 1591-1604. https://doi.org/10.1007/s11357-021-00365-7
|
| [45] |
Berry BJ, Mjelde E, Carreno F, et al. (2023) Preservation of mitochondrial membrane potential is necessary for lifespan extension from dietary restriction. GeroScience 45: 1573-1581. https://doi.org/10.1007/s11357-023-00766-w
|
| [46] |
Sutherland TC, Sefiani A, Horvat D, et al. (2021) Age-dependent decline in neuron growth potential and mitochondria functions in cortical neurons. Cells 10: 1625. https://doi.org/10.3390/cells10071625
|
| [47] |
Zhang H, Alder NN, Wang W, et al. (2020) Reduction of elevated proton leak rejuvenates mitochondria in the aged cardiomyocyte. Elife 9: e60827. https://doi.org/10.7554/eLife.60827
|
| [48] |
Barnstable CJ, Zhang M, Tombran-Tink J (2022) Uncoupling proteins as therapeutic targets for neurodegenerative diseases. Int J Mol Sci 23: 5672. https://doi.org/10.3390/ijms23105672
|































Figures(32) / Tables(1)

Mahmoud Abdallat, Abdallah Barjas Qaswal, Majed Eftaiha, Abdel Rahman Qamar, Qusai Alnajjar, Rawand Sallam, Lara Kollab, Mohammad Masa'deh, Anas Amayreh, Hiba Mihyar, Hesham Aboushakra, Bayan Alkelani, Rawan Owaimer, Mohannad Abd-Alhadi, Salwa Ireiqat, Fahed Turk, Ahmad Daoud, Bashar Darawsheh, Ahmad Hiasat, Majd Alhalaki, Shahem Abdallat, Salsabiela Bani Hamad, Rand Murshidi. A mathematical modeling of the mitochondrial proton leak via quantum tunneling[J]. AIMS Biophysics, 2024, 11(2): 189-233. doi: 10.3934/biophy.2024012







 DownLoad:
DownLoad: