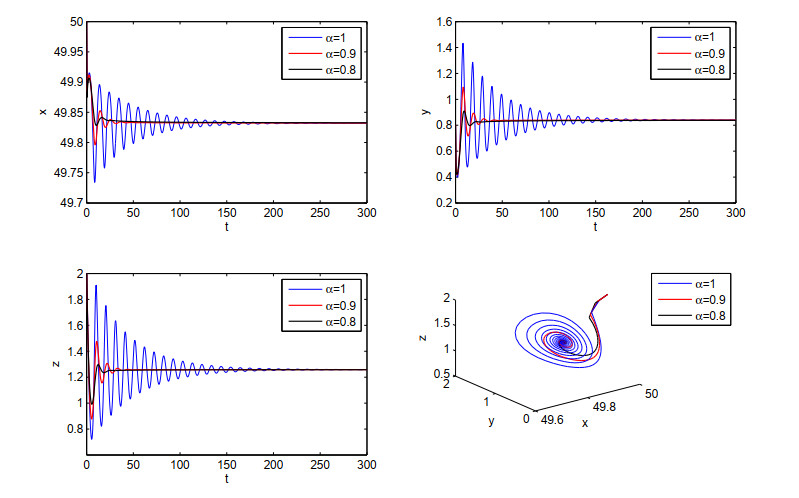
Figure 1.
Stability of the equilibrium E∗ for α=1, α=0.9 and α=0.8.





Citation: Alessandro Venditti, Claudio Frezza, Giacomo Rossi, Mirella Di Cecco, Giampiero Ciaschetti, Mauro Serafini, Armandodoriano Bianco. Secondary metabolites with ecologic and medicinal implications in Anthemis cretica subsp. petraea from Majella National Park[J]. AIMS Molecular Science, 2016, 3(4): 648-660. doi: 10.3934/molsci.2016.4.648
| [1] | Peter A. Braza . A dominant predator, a predator, and a prey. Mathematical Biosciences and Engineering, 2008, 5(1): 61-73. doi: 10.3934/mbe.2008.5.61 |
| [2] | Jian Zu, Wendi Wang, Bo Zu . Evolutionary dynamics of prey-predator systems with Holling type II functional response. Mathematical Biosciences and Engineering, 2007, 4(2): 221-237. doi: 10.3934/mbe.2007.4.221 |
| [3] | Yongli Cai, Malay Banerjee, Yun Kang, Weiming Wang . Spatiotemporal complexity in a predator--prey model with weak Allee effects. Mathematical Biosciences and Engineering, 2014, 11(6): 1247-1274. doi: 10.3934/mbe.2014.11.1247 |
| [4] | Jocirei D. Ferreira, Wilmer L. Molina, Jhon J. Perez, Aida P. González . Stability and bifurcation analysis in predator-prey system involving Holling type-II functional response. Mathematical Biosciences and Engineering, 2025, 22(10): 2559-2594. doi: 10.3934/mbe.2025094 |
| [5] | Xiaoyuan Chang, Junjie Wei . Stability and Hopf bifurcation in a diffusivepredator-prey system incorporating a prey refuge. Mathematical Biosciences and Engineering, 2013, 10(4): 979-996. doi: 10.3934/mbe.2013.10.979 |
| [6] | Yajie Sun, Ming Zhao, Yunfei Du . Multiple bifurcations of a discrete modified Leslie-Gower predator-prey model. Mathematical Biosciences and Engineering, 2023, 20(12): 20437-20467. doi: 10.3934/mbe.2023904 |
| [7] | Eric M. Takyi, Charles Ohanian, Margaret Cathcart, Nihal Kumar . Dynamical analysis of a predator-prey system with prey vigilance and hunting cooperation in predators. Mathematical Biosciences and Engineering, 2024, 21(2): 2768-2786. doi: 10.3934/mbe.2024123 |
| [8] | Andrés Sanchéz, Leon A. Valencia, Jorge M. Ramirez Osorio . The Stochastic Gause Predator-Prey model: Noise-induced extinctions and invariance. Mathematical Biosciences and Engineering, 2025, 22(8): 1999-2019. doi: 10.3934/mbe.2025073 |
| [9] | Jian Zu, Wendi Wang, Bo Zu . Letter to the editors. Mathematical Biosciences and Engineering, 2007, 4(4): 755-755. doi: 10.3934/mbe.2007.4.755 |
| [10] | Christopher M. Kribs-Zaleta . Sharpness of saturation in harvesting and predation. Mathematical Biosciences and Engineering, 2009, 6(4): 719-742. doi: 10.3934/mbe.2009.6.719 |
Population dynamics has always been an important research object of biomathematics. Various groups often have complex interspecific relationships, such as predation, competition, parasitism and mutualism [1]. Predation behavior, as a widespread interspecies relationship, has been widely studied. Lotka and Volterra were the first to propose a predator-prey system to describe the widespread interspecies relationship of predation [2]. Holling further proposed three functional responses of predators to describe the energy transfer between predators and prey [3]. These three functional responses have been applied and perfected by many scientists [4]. In 1991, Hastings and Powell proposed a food chain system with chaotic dynamics and studied the dynamics of the model [5]. In recent years, many mathematicians have also studied the development and improvement of Hastings-Powell food chain models [6,7,8,9].
Fractional calculus is a generalization of traditional calculus, and its order can be composed of integers, fractions or complex numbers[10]. Fractional calculus can better describe some systems or processes with memory and hereditary properties, and it has been widely used in many fields, such as physics, secure communication, system control, neural networks, and chaos[11,12]. The method of solving the fractional model has also been widely studied[13,14]. In [15], the Caputo fractional derivative operator is used instead of the integer first derivative to establish an effective numerical method for solving the dynamics of the reaction-diffusion model based on a new implicit finite difference scheme. In [16], a numerical approximation for the Caputo-Fabrizio derivative is used to study the dynamic complexity of a predator-prey system with a Holling-type functional response. In [17], a new fractional chaotic system described by the Caputo fractional derivative is presented, and how to use the bifurcation diagram of this chaotic system to detect chaotic regions is analyzed. In [18], the generalization of Lyapunov's direct method applying Bihari's and Bellman-Gronwall's inequalities to Caputo-type fractional-order nonlinear systems is proposed. In [19], the Fourier spectral method is introduced to explore the dynamic richness of two-dimensional and three-dimensional fractional reaction-diffusion equations. In [20], the spatial pattern formation of the predator-prey model with different functional responses was studied. [21] studied the numerical solution of the space-time fractional reaction-diffusion problem that simulates the dynamic and complex phenomena of abnormal diffusion.
Since most biological mathematical models have long-term memory, fractional differential equations can more accurately and reliably describe the actual dynamic process [1,22]. [23] proposed fractional predator-prey models and fractional rabies models and studied their equilibrium points, stability and numerical solutions. In [24], the authors studied the stability of a fractional-order system by the Lyapunov direct method, which substantially developed techniques to study the stability of fractional-order population models. In [9], the authors extended the Hastings-Powell food chain system to fractional order and analyzed its dynamic behavior.
As an important research object of biological mathematics and control theory, population model control has received extensive research and development in recent years [25,26,27]. In [28], the authors conducted random detection and contacted tracking on the HIV/AIDS epidemic model and used the Adams-type predictor-corrector method to perform fractional optimal control of the model, which significantly reduced the number of AIDS patients and HIV-infected patients. In [29], the authors applied the time-delay feedback controller to the fractional-order competitive Internet model to solve the bifurcation control problem of this model. In [30], the author considered the influence of additional predators on the Hastings-Powell food chain model and studied the control of chaos in this model.
Biological models are widely studied by scientists, but many classic models study food chain models composed of herbivores and carnivores, and omnivores are rarely considered. In fact, omnivores are widespread in nature and play an important role in the food chain. In this article, the existence of omnivores is fully considered, and a food chain model in which herbivores, omnivores and carnivores coexist is studied. Based on these works, this paper proposes a fractional food chain model with a Holling type-II functional response. The main contributions of this paper are as follows. First, this paper proves the existence and uniqueness of the solution and the nonnegativity and boundedness of the solution. Second, the equilibrium point of the model is calculated, and the local stability of the equilibrium point is proven. Third, a controller is designed to prove the global asymptotic stability of the system by using the Lyapunov method.
This paper is organized as follows. In Section 2, the definitions and lemmas are given, and the food chain model is established. In Section 3, the existence, uniqueness, nonnegativity and boundedness are proven, and the local stability of the equilibrium point of the model is studied. The global stability of the model is studied through the controller. In Section 4, numerical simulations are performed to verify the theoretical results. The conclusion of this article is given in Section 5.
In this section, some basic knowledge about fractional equations and the theorems and lemmas used in this paper are given, and the fractional food chain system is introduced.
Definition 1. [10]. The Caputo fractional derivative of order α of a function f, R+→R, is defined by
| Dαtf(t)=1Γ(n−α)∫t0fn(τ)(t−τ)α+1−ndτ,(n−1<α<n), n∈Z+, |
where Γ(⋅) is the Gamma function. When 0<α<1,
| Dαtf(t)=1Γ(1−α)∫t0f′(τ)(t−τ)αdτ. |
Definition 2. [31]. When the order a>0, for a function f:(0,∞)→R, the Riemann-Liouville representation of the fractional integral operator is defined by
| RLD−atf(t)=RLIatf(t)=1Γ(a)∫t0(t−τ)a−1f(τ)dτ,t>0, |
| RLI0tg(t)=g(t), |
where a>0 and Γ(⋅) is the Gamma function.
Lemma 1. (Generalized Gronwall inequality) [32]. Assume that m≥0, γ>0, and a(t) are nonnegative, locally integrable, and nondecreasing functions defined on 0≤t≤T(T≤∞). In addition, h(t) is a nonnegative, locally integrable function defined in 0≤t≤T and satisfies
| h(t)≤a(t)+m∫t0(t−s)γ−1h(t)ds, |
then,
| h(t)=a(t)Eγ(mΓ(γ)tγ), |
where the Mittag-Leffler function Eγ(z)=∞∑k=0zkΓ(kγ+1).
Lemma 2. [10]. Consider the fractional-order system
| {Dαtx(t)=f(t,x(t)),x(0)=x0, | (2.1) |
where f(t,x(t)) defined in R+×Rn→Rn and α∈(0,1].
The local asymptotic stability of the equilibrium point of this system can be derived from |arg(λi)|>απ2, where λi are the eigenvalues of the Jacobian matrix at the equilibrium points.
Lemma 3. [24]. Consider the system
| {Dαtx(t)=f(t,x(t)),x(0)=xt0, | (2.2) |
where α∈(0,1], f:[t0,∞)×Ω→Rn, and Ω∈Rn; if f(t,x) satisfies the local Lipschitz condition about x on [t0,∞)×Ω, then there exists a unique solution of (2.2).
Lemma 4. [33]. The function x(t)∈R+ is continuous and derivable; then, for any t≥t0
| Dαt[x(t)−x∗−x∗lnx(t)x∗]≤(1−x∗x(t))Dαtx(t),x∗∈R+,∀α∈(0,1). |
There are a variety of complex biological relationships in nature. Predation is the most important biological relationship, and it has received attention from and been studied by many scientists. In [5], the author proposed a three-species food chain model. The model consists of one prey ˆX and two predators ˆY and ˆZ. The top predator ˆZ feeds on the secondary predator ˆY, and the secondary predator ˆY feeds on the prey ˆX. This is the famous Hastings-Powell model:
| {dˆXdT=ˆRˆX(1−ˆXˆK)−^C1^A1ˆXˆY^B1+ˆX,dˆYdT=^A1ˆXˆY^B1+ˆX−^A2ˆYˆZ^B2+ˆY−^D1ˆY,dˆZdT=^C2^A2ˆYˆZ^B2+ˆY−^D2ˆZ, | (2.3) |
where ˆR and ˆK represent the intrinsic growth rates and environmental carrying capacity, respectively. For i=1,2, parameters^Ai, ^Bi, ^Ci and ^Di are the predation coefficients, half-saturation constant, food conversion coefficients and death rates.
The Hastings-Powell model considers a food chain composed of herbivores, small carnivores and large carnivores but does not consider the existence of omnivores. We consider a food chain consisting of small herbivores X, medium omnivores Y and large carnivores Z. Among them, omnivores Y prey on herbivores X, and carnivores Z prey on omnivores Y. They all respond according to Holling II type. This system can be expressed mathematically as
| {dXdT=R1X(1−XK1)−C1A1XYB1+X,dYdT=R2Y(1−YK2)+A1XYB1+X−A2YZB2+Y,dZdT=C2A2YZB2+Y−DZ, | (2.4) |
where X, Y and Z represent the densities of the prey population, primary predator population and top-predator population, respectively. For i=1,2, parameters Ri, Ki, Ai, Bi and Ci are the intrinsic growth rates, environmental carrying capacity, predation coefficients, half-saturation constant and food conversion coefficients, respectively. The parameter D is the death rates for Z.
Then, we obtain the following dimensionless version of the food chain model:
| {dxdt=r1x(1−xK1)−a1xy1+b1x,dydt=r2y(1−yK2)+a1xy1+b1x−a2yz1+b2y,dzdt=a2yz1+b2y−dz, | (2.5) |
the independent variables x, y and z are dimensionless population variables; t represents a dimensionless time variable; and ai, bi(i=1,2) and d are positive.
Research results show that using fractional derivatives to model real-life biological problems is more accurate than classical derivatives[15]. To better analyze the dynamics between these three populations, we studied the following fractional-order Hastings-Powell System food chain model:
| {Dαtx=r1x(1−xK1)−a1xy1+b1x,Dαty=r2y(1−yK2)+a1xy1+b1x−a2yz1+b2y,Dαtz=a2yz1+b2y−dz, | (2.6) |
where α∈(0,1) is the fractional order.
Theorem 1. The fractional-order Hastings-Powell System food chain model (2.6) has a unique solution.
Proof: We will study the existence and uniqueness of the system (2.6) in [0,T]×Ω, where Ω={(x,y,z)∈R3:0≤x,y,z≤H}. Let S=(x,y,z), ˉS=(ˉx,ˉy,ˉz), F(S)=(F1(S),F2(S),F3(S)) and
| {F1(S)=Dαtx=r1x(1−xK1)−a1xy1+b1x,F2(S)=Dαty=r2y(1−yK2)+a1xy1+b1x−a2yz1+b2y,F3(S)=Dαtz=a2yz1+b2y−dz, | (3.1) |
For any S,ˉS∈Ω, it follows from (3.1) that
| ‖F(S)−F(ˉS)‖=|F1(S)−F1(ˉS)|+|F2(S)−F2(ˉS)|+|F3(S)−F3(ˉS)|=|r1x(1−xK1)−a1xy1+b1x−(r1ˉx(1−ˉxK1)−a1ˉxˉy1+b1ˉx)|+|r2y(1−yK2)+a1xy1+b1x−a2yz1+b2y−(r2ˉy(1−ˉyK2)+a1ˉxˉy1+b1ˉx−a2ˉyˉz1+b2ˉy)|+|a2yz1+b2y−dz−(a2ˉyˉz1+b2ˉy−dˉz)|≤r1|x(1−xK1)−ˉx(1−ˉxK1)|+r2|y(1−yK2)−ˉy(1−ˉyK2)|+2a1|xy1+b1x−ˉxˉy1+b1ˉx|+2a2|yz1+b2y−ˉyˉz1+b2ˉy|+d|z−ˉz|≤r1|x−ˉx|+r2|y−ˉy|+r1K1|x2−ˉx2|+r2K2|y2−ˉy2|+d|z−ˉz|+|xy−ˉxˉy+b1ˉxxy−b1ˉxxˉy(1+b1x)(1+b1ˉx)|+|yz−ˉyˉz+b2ˉyyz−b2ˉyyˉz(1+b2y)(1+b2ˉy)|≤r1|x−ˉx|+r2|y−ˉy|+r1K1|(x+ˉx)(x−ˉx)|+r2K2|(y+ˉy)(y−ˉy)|+d|z−ˉz|+|xy−ˉxˉy+b1xˉx(y−ˉy)|+|yz−ˉyˉz+b2yˉy(z−ˉz)|≤r1|x−ˉx|+r2|y−ˉy|+r1MK1|(x−ˉx)|+r2MK2|(y−ˉy)|+d|z−ˉz|+|xy−xˉy+xˉy−ˉxˉy|+b1|xˉx||y−ˉy|+|yz−yˉz+yˉz−ˉyˉz|+b2|yˉy||z−ˉz|≤(r1+r1MK1)|x−ˉx|+(r2+r2MK2)|y−ˉy|+d|z−ˉz|+M|y−ˉy|+M|x−ˉx|+b1M2|y−ˉy|+M|z−ˉz|+M|y−ˉy|+b2M2|z−ˉz|=(r1+r1MK1+M)|x−ˉx|+(r2+r2MK2+2M+b1M2)|y−ˉy|+(d+M+b2M2)|z−ˉz|≤L‖S−ˉS‖. |
where L=max{r1+r1MK1+M,r2+r2MK2+2M+b1M2,d+M+b2M2}, Based on Lemma 3, F(S) satisfies the Lipschitz condition with respect to S in Ω. According to the Banach fixed point theorem in [34], system (2.6) has a unique solution in Ω.
Theorem 2. Set A={x,y,z)∈R3:0<x+y+z<K1(r1+v)24r1v+K2(r2+v)24r2v} as a positively invariant set of system (2.6), and the solutions are bounded.
Proof: let g(t)≜g(x(t),y(t),z(t))=x(t)+y(t)+z(t),
| Dαtg(t)+vg(t)=(r1+v)x(t)−r1K1x2(t)+(r2+v)y(t)−r2K2y2(t)−(d−v)z(t)=−r1K1(x(t)−K1(r1+v)2r1)2−r2K2(y(t)−K2(r2+v)2r2)2−(d−v)z(t)+K1(r1+v)24r1+K2(r2+v)24r2, |
let
| u=K1(r1+v)24r1+K2(r2+v)24r2,v=d. |
According to the positive knowledge of all parameters and the nonnegativity of the solutions,
| Dαtg(t)+vg(t)≤u, |
we obtain
| g(t)≤uv+[g(0)−uv]Eα(−vtα), |
Since Eα(−vtα)≥0, when g(0)≤uv, limt→∞supg(t)≤uv. According to the nonnegativity of the system (2.6), g(t)≥0(∀t≥0); hence, A={x,y,z)∈R3:0<x+y+z<K1(r1+v)24r1v+K2(r2+v)24r2v} is a positively invariant set of system (2.6), and the solutions are bounded.
Let
| {r1x(1−xK1)−a1xy1+b1x=0,r2y(1−yK2)+a1xy1+b1x−a2yz1+b2y=0,a2yz1+b2y−dz=0, |
Then, the equilibrium points are E0=(0,0,0), E1=(K1,0,0), E2=(0,K2,0) and E∗=(x∗,y∗,z∗), where
| x∗=b1K1−12b1+√b21K21r21+2b1K1r21−4a1b1K1r1y∗+y2∗2b1r1,y∗=da2−b2d,z∗=a1x∗(b2y∗+1)a2(b1x∗+1)−r2(b2y∗+1)(y∗−K2)a2K2. |
For system (2.6), the Jacobian matrix at the equilibrium point (x′,y′,z′) is
| J(x′,y′,z′)=[r1−2r1x′K1−a1de1e23−e50e4r1+e5−a2z′e2−2dr2K2e1+a2b2dz′e1e22−d0e21z′a20], |
where e1=a2−b2d, e2=b2de1+1, e3=1+b1x′, e4=2r1x′K1+a1de1e23, e5=a1x′e3.
Theorem 3.3. For system (2.6), the equilibrium points E0 and E1 are saddle points.
Proof: The Jacobian matrices evaluated at E0 and E1 are
| J(0,0,0)=[r1000r2000−d],J(K1,0,0)=[−r1−a1K1b1K1+100r2+a1K1b1K1+1000−d], |
According to Lemma 2, when the eigenvalues are all real numbers and all negative, the equilibrium points are locally asymptotically stable.
The eigenvalues of J(E0) are λ01=r1, λ02=r2 and λ03=−d. The eigenvalues of J(E1) are λ11=−r1, λ12=r2+a1K1b1K1+1 and λ13=−d.
Then, we have λ01,λ02>0, λ03,λ11<0, and λ13<0, so the equilibrium points E0 and E1 are saddle points.
Theorem 4. For system (2.6), if r1<a1K2 and a2K2(b2K2+1)<d, then the equilibrium point E2=(0,K2,0) is locally asymptotically stable.
Proof: The Jacobian matrix evaluated at E2 is
| J(0,K2,0)=[r1−a1K200a1K2−r2−a2K2b2K2+100a2K2b2K2+1−d], |
The eigenvalues of J(E2) are λ1=r1−a1K2, λ2=−r2 and λ3=a2K2b2K2+1−d.
Therefore, if r1<a1K2 and a2K2(b2K2+1)<d, the equilibrium point E2 is locally asymptotically stable.
The characteristic equation of equilibrium points E∗=(x∗,y∗,z∗) is given as
| P(λ)=λ3+Aλ2+Bλ+C=0, | (3.2) |
where
A=2f4−2r1−e5+f2z∗+f1de23+f5−f2b2dz∗e1e2,
B=(B1+B2),
B1=e4e5+e5r1+r21−2r1f4−f2r1z∗−2e5f4+e21dz∗a2−f6e5−f6r1−2r1f5,
B2=2e23f1f5+2f2f4z∗+a1f3e23z∗+4dr2f4e1k1+r1f7−f1f7de3−2f4f7,
C=dz∗(a1dK1e23−e1K1r1+2e1r1x∗)f1K1,
where f1=a1e1, f2=a2e2, f3=f2de1, f4=r1x∗K1, f5=r2de1K2, f6=a1de23e1, f7=b2f3z∗e2.
For Eq. (3.2), define the discriminant as
| D(P)=18ABC+(AB)2−4CA2−4B3−27C2, |
With reference to the results of [35] and [36], we obtain the following fractional Routh-Hurwitz conditions:
1. If D(P)>0, A>0, C>0, and AB−C>0, E∗ is locally asymptotically stable.
2. If D(P)<0 and A≥0, B≥0, and C>0, when α<23, E∗ is locally asymptotically stable.
3. If D(P)<0, A>0, B>0, and AB=C, then for all α∈(0,1), E∗ is locally asymptotically stable.
Theorem 5. If D(P)<0, C>0 and AB≠C, then α∗∈(0,1) exists; when α∈(0,α∗), E∗ is locally asymptotically stable; when α∈(α∗,1), E∗ is unstable. The system diverges at the critical value E∗.
Proof: If D(P)<0, then the eigenvalues of Eq. (3.2) have one real root λ1=a and two complex conjugate roots λ2,3=b±ci. Then, Eq. (3.2) can be written as
| P(λ)=(λ−a)[λ−(b+ci)][λ−(b−ci)]=0, | (3.3) |
where A=−a−2b, B=b2+c2+2ab, C=−a(b2+c2), c>0, a,b,c∈R.
From C>0, then a<0, and then |arg(λ1)|=π>απ2.
From AB≠C, then −a2b+b(b2+c2)≠−2ab2 ⟹ −2b[(a+b)2+c2]≠0 ⟹ b≠0 and (a+b)2+c2≠0.
Thus, we can obtain |arg(λ2,3)|=|arctan(cb)|=arctan|cb|∈(0,π2).
Then, α∗∈(0,1) exists; when α∈(0,α∗), απ2<arctan|cb|, according to Lemma 2, E∗ is locally asymptotically stable, and when α∈(α∗,1), απ2>arctan|cb|, E∗ is unstable.
To study the asymptotic stability of system (2.6), three controllers will be added. The controller is proposed as follows: μ1=m1x(x−x∗), μ2=m2y(y−y∗), and μ3=m3z(z−z∗). where m1, m2 and m3 represent negative feedback gains, which are defined as real numbers. Clearly, if mi=0(i=1,2,3) or x=x∗(y=y∗,z=z∗), then μi=0(i=1,2,3), so it will not change the equilibrium point of system (2.6).
Controllers added into system (2.6) as follows
| {Dαtx=r1x(1−xK1)−a1xy1+b1x−m1x(x−x∗),Dαty=r2y(1−yK2)+a1xy1+b1x−a2yz1+b2y−m2y(y−y∗),Dαtz=a2yz1+b2y−dz−m3z(z−z∗), | (3.4) |
One gives a Lyapunov function as:
| V(x,y,z)=x−x∗−x∗lnxx∗+y−y∗−y∗lnyy∗+z−z∗−z∗lnzz∗. |
then,
| DαtV≤x−x∗xDαx+y−y∗yDαy+z−z∗zDαz=(x−x∗)(r1−r1xK1−a1y1+b1x)−m1(x−x∗)2+(y−y∗)(r2−r2yK2+a1x1+b1x−a2z1+b2y)−m2(y−y∗)2+(z−z∗)(a2y1+b2y−d)−m3(z−z∗)2. |
Consider E∗ to be the equilibrium point:
| {r1−r1x∗K1−a1y∗1+b1x∗=0,r2−r2y∗K2+a1x∗1+b1x∗−a2z∗1+b2y∗=0,a2y∗1+b2y∗−d=0, |
According to Lemma 4, we can obtain
| DαtV≤(x−x∗)(r1x∗K1+a1y∗1+b1x∗−r1xK1−a1y1+b1x)−m1(x−x∗)2+(y−y∗)(r2y∗K2−a1x∗1+b1x∗+a2z∗1+b2y∗−r2yK2+a1x1+b1x−a2z1+b2y)−m2(y−y∗)2+(z−z∗)(a2y1+b2y−a2y∗1+b2y∗)−m3(z−z∗)2=a1(x−x∗)(y∗1+b1x∗−y1+b1x)+a1(y−y∗)(x1+b1x−x∗1+b1x∗)+a2(y−y∗)(z∗1+b2y∗−z1+b2y)+a2(z−z∗)(y1+b2y−y∗1+b2y∗)−(m1+r1K1)(x−x∗)2−(m2+r2K2)(y−y∗)2−m3(z−z∗)2=a1(x−x∗)(y∗1+b1x∗−y∗1+b1x+y∗1+b1x−y1+b1x)+a1(y−y∗)(x1+b1x−x∗1+b1x+x∗1+b1x−x∗1+b1x∗)+a2(y−y∗)(z∗1+b2y∗−z∗1+b2y+z∗1+b2y−z1+b2y)+a2(z−z∗)(y1+b2y−y∗1+b2y+y∗1+b2y−y∗1+b2y∗)−(m1+r1K1)(x−x∗)2−(m2+r2K2)(y−y∗)2−m3(z−z∗)2=a1(x−x∗)(b1y∗(x−x∗)(1+b1x∗)(1+b1x)+y∗−y1+b1x)+a1(y−y∗)(x−x∗1+b1x+b1x∗(x∗−x)(1+b1x∗)(1+b1x))+a2(y−y∗)(b2z∗(y−y∗)(1+b2y∗)(1+b2y)+z∗−z1+b2y)+a2(z−z∗)(y−y∗1+b2y+b2y∗(y∗−y)(1+b2y∗)(1+b2y))−(m1+r1K1)(x−x∗)2−(m2+r2K2)(y−y∗)2−m3(z−z∗)2≤a1b1y∗1+b1x∗(x−x∗)2+a2b2z∗1+b2y∗(y−y∗)2+a1b1x∗1+b1x∗(x−x∗)(y∗−y)+a2b2y∗1+b2y∗(y−y∗)(z∗−z)−(m1+r1K1)(x−x∗)2−(m2+r2K2)(y−y∗)2−m3(z−z∗)2≤a1b1x∗2(1+b1x∗)((x−x∗)2+(y−y∗)2)+a2b2y∗2(1+b2y∗)((y−y∗)2+(z−z∗)2)+(a1b1y∗1+b1x∗−m1−r1K1)(x−x∗)2+(a2b2z∗1+b2y∗−m2−r2K2)(y−y∗)2−m3(z−z∗)2=(a1b1(2y∗+x∗)2(1+b1x∗)−m1−r1K1)(x−x∗)2+(a2b2y∗2(1+b2y∗)−m3)(z−z∗)2+(a2b2(2z∗+y∗)2(1+b2y∗)+a1b1x∗2(1+b1x∗)−m2−r2K2)(y−y∗)2. |
When m1≥a1b1(2y∗+x∗)2(1+b1x∗)−r1K1, m2≥a2b2(2z∗+y∗)2(1+b2y∗)+a1b1x∗2(1+b1x∗)−r2K2, and m3≥a2b2y∗2(1+b2y∗), it follows that DαV≤0. We can show that the equilibrium point E∗ is uniformly asymptotically stable.
In this section, we use the Adams-Bashforth-Molton predictor-corrector algorithm numerical simulation. This method is described in detail in [37] and [38].
Example 1. In system (2.6), let r1=1, r2=0.6, K1=50, K2=10, a1=1, a2=0.6, b1=5, b2=0.01 and d=0.5. System (2.6) has a positive equilibrium point E∗=(49.8479,0.8403,1.2597). According to Theorem 3.5, when α=1, α=0.9, α=0.8, and E∗ is locally asymptotically stable, it can be seen from Figure 1 that the order α will affect the speed at which the system converges to the equilibrium point. The relevant results are shown in Figure 1.
Example 2. In system (2.6), let r1=1, r2=0.6, K1=50, K2=30, a1=1, a2=0.6, b1=5, b2=0.2 and d=0.2. System (2.6) has a positive equilibrium point E∗=(49.9382,0.3571,1.4158). It follows from Theorem 3.5 that system (2.6) has a bifurcation at α∗. When α=0.95 and α=0.8, E∗ is locally asymptotically stable, and when α=0.98, E∗ is unstable. The relevant results are shown in Figure 2.

Example 3. To verify the sensitivity of the system (2.6) to initial conditions and other parameters, according to the method in [39], apply the positive Euler format to transform the differential model into the following discrete form:
| {xt+1=xt+δ(r1xt(1−xtK1)−a1xtyt1+b1xt),yt+1=yt+δ(r2yt(1−ytK2)+a1xtyt1+b1xt−a2ytzt1+b2yt),zt+1=zt+δ(a2ytzt1+b2yt−dzt), | (4.1) |
where δ is the time step size. We use the parameters of Example 1 to study Lyapunov exponents. Figure 3 shows that system (2.6) is in a stable state and is less sensitive to initial conditions.

This paper studies a new fractional-order food chain model with a Holling type-II functional response. First, the existence, uniqueness, nonnegativity and boundedness of the solution of the model are discussed. Second, the local stability of each equilibrium point is discussed. Third, controllers μ1=m1x(x−x∗), μ2=m2y(y−y∗) and μ3=m3z(z−z∗) are proposed and added to the system. Using the Lyapunov method, sufficient conditions for the positive equilibrium point to reach the global uniformly asymptotically stable state are obtained. Finally, we use numerical simulations to verify the theoretical results.
This work was supported by the Shandong University of Science and Technology Research Fund (2018 TDJH101).
The author declares no conflicts of interest in this paper.
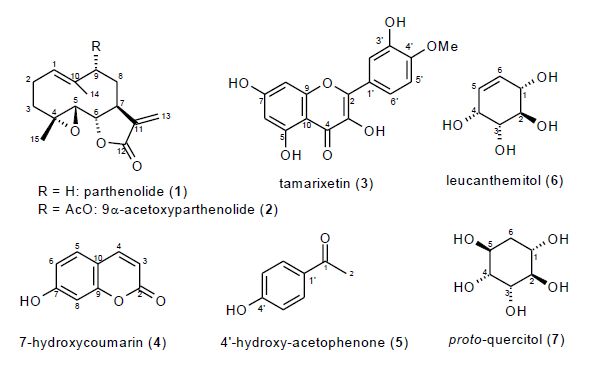
| [1] | Baytop T (1999) Türkiye’de Bitkiler ile Tedavi, Geçmis¸te ve Bugün (Therapy with Medicinal Plants in Turkey, Past and Present), 2nd ed., Nobel Tıp Kitabevleri, Istanbul, Turkey. |
| [2] |
Kultur S (2007) Medicinal plants used in Kırklareli Province (Turkey). J Ethnopharmacol 111: 341-364. doi: 10.1016/j.jep.2006.11.035

|
| [3] |
Ugurlu E, Secmen O (2008) Medicinal plants popularly used in the villages of Yunt Mountain (Manisa-Turkey). Fitoterapia 79: 126-131. doi: 10.1016/j.fitote.2007.07.016
|
| [4] |
Gonenc T, Argyropoulou C, Erdogan T, et al. (2011) Chemical constituents from Anthemis wiedemanniana Fisch. & Mey. Biochem Syst Ecol 39: 51-55. doi: 10.1016/j.bse.2011.01.007
|
| [5] |
Pavlovic M, Kovacevic N, Couladis M, et al. (2006) Phenolic constituents of Anthemis triumfetti (L.) DC. Biochem Syst Ecol 34: 449-452. doi: 10.1016/j.bse.2005.11.009
|
| [6] |
Kivcak B, Mert T, Saglam H, et al. (2007) Chemical composition and antimicrobial activity of the essential oil of Anthemis wiedemanniana from Turkey. Chem Nat Compd 43: 47-51. doi: 10.1007/s10600-007-0029-x
|
| [7] |
Uzel A, Guvensen A, Cetin E (2004) Chemical composition and antimicrobial activity of the essential oils of Anthemis xylopoda O. Schwarz from Turkey. J Ethnopharm 95: 151-154. doi: 10.1016/j.jep.2004.06.034
|
| [8] |
Javidnia K, Miri R, Kamalinejad M, et al. (2004) Chemical composition of the essential oils of Anthemis altissima L. grown in Iran. Flavour Fragr J 19: 213-216. doi: 10.1002/ffj.1277
|
| [9] |
Bruno M, Bonoi ML, Vassallo N, et al. (1997) Guaianolides and other terpenoids from Anthemis aetnensis. Phytochemistry 45: 375-377. doi: 10.1016/S0031-9422(96)00846-1
|
| [10] | Hofer O, Greger H (1985) New Sesquiterpene-Coumarin Ethers from Anthemis cretica. Liebigs Annalen der Chemie 6: 1136-1144. |
| [11] |
Vajs V, Bulatovic V, Fodulovic-Savikin K, et al. (1999) Highly oxygenated guaianolides from Anthemis cretica subsp. cretica. Phytochemistry 50: 287-291. doi: 10.1016/S0031-9422(98)00504-4
|
| [12] | Tenore M (1811-1815) Flora Neapolitana ossia descrizione delle piante indigene del Regno di Napoli e delle più rare specie di piante esotiche coltivate nei giardini, Vol.1. Stamperia reale, Tip. del Giornale Enciclopedico e Stamperia Francese, Napoli. |
| [13] |
Peruzzi L, Conti F, Bartolucci F (2014) An inventory of vascular plants endemic to Italy. Phytotaxa 168: 1-75. doi: 10.11646/phytotaxa.168.1.1
|
| [14] | Conti F (1998) An annotated checklist of the flora of the Abruzzo. Bocconea 10: 276. |
| [15] |
Venditti A, Serrilli AM, Di Cecco M, et al. (2013) Phytochemical composition of polar fraction of Stachys germanica L. subsp. salviifolia (Ten.) Gams, a typical plant of Majella National Park. Nat Prod Res 27: 190-193. doi: 10.1080/14786419.2012.661732
|
| [16] |
Venditti A, Serrilli AM, Di Cecco M, et al. (2012) Phytochemical analysis of Plantago sempervirens from Majella National Park. Nat Prod Res 26: 2035-2039. doi: 10.1080/14786419.2011.633520
|
| [17] |
Venditti A, Maggi F, Vittori S, et al. (2014) Volatile compounds from Achillea tenorii (Grande) growing in the Majella National Park (Italy). Nat Prod Res 28: 1699-1704. doi: 10.1080/14786419.2014.940349
|
| [18] |
Venditti A, Maggi F, Vittori S, et al. (2015) Antioxidant and α-glucosidase inhibitory activities of Achillea tenorii. Pharm Biol 53: 1505-1510. doi: 10.3109/13880209.2014.991833
|
| [19] |
Tiuman TS, Ueda-Nakamura T, Cortez DAG, et al. (2005) Antileishmanial activity of parthenolide, a sesquiterpene lactone isolated from Tanacetum parthenium. Antimicrob. Agents Chemother 49: 176-182. doi: 10.1128/AAC.49.11.176-182.2005
|
| [20] |
Worawalai W, Rattanangkool E, Vanitcha A, et al. (2012) Concise synthesis of (+)-conduritol F and inositol analogues from naturally available (+)-proto-quercitol and their glucosidase inhibitory activity. Bioorg Med Chem Lett 22: 1538-1540. doi: 10.1016/j.bmcl.2012.01.007
|
| [21] |
Bohlmann F, Zdero C (1975) Naturally occurring terpene derivatives. XLVI. A new sesquiterpene lactone from Matricaria suffructicosa var. leptoloba. Chem Ber 108: 437-439. doi: 10.1002/cber.19751080208
|
| [22] |
Bruno M, Diaz JG, Herz W (1991) Germacranolides from Anthemis cupaniana. Phytochemistry 30: 3458-3460. doi: 10.1016/0031-9422(91)83229-E
|
| [23] |
Lewin G, Maciuk A, Thoret S, et al. (2010) Semisynthesis of Natural Flavones Inhibiting Tubulin Polymerization, from Hesperidin. J Nat Prod 73: 702-706. doi: 10.1021/np100065v
|
| [24] |
Tzenge‐Lien S, Ya‐Ling L (2005) Epoxidation of Protected (1,4,5)-Cyclohex-2-ene-triols and Their Acid Hydrolysis to Synthesize Quercitols from D-(-)-Quinic Acid. Synth Commun 35: 1809-1817. doi: 10.1081/SCC-200063960
|
| [25] |
Dhanuskodi S, Manikandan S (2005) EPR investigations on γ-irradiated 4-hydroxyacetophenone single crystals. An NLO material. Radiat Eff Defects Solids 160: 197-205. doi: 10.1080/10420150500275569
|
| [26] |
Timonen JM, Nieminen RM, Sareila O, et al. (2011) Synthesis and anti-inflammatory effects of a series of novel 7-hydroxycoumarin derivatives. Eur J Med Chem 46: 3845-3850. doi: 10.1016/j.ejmech.2011.05.052
|
| [27] | Bianco A, Serrilli AM, Venditti A, et al. (2016) Endemic Plants of Italy and Their Peculiar Molecular Pattern. In: Studies in Natural Products Chemistry (Bioactive Natural Products), Ed. Atta-Ur-Rahman, Elsevier Science Publishers - Amsterdam. Vol. 50, Chap. 7, pp. 215-247. |
| [28] |
Staneva JD, Todorova MN, Evstatieva LN (2008) Sesquiterpene lactones as chemotaxonomic markers in genus Anthemis. Phytochemistry 69: 607-618. doi: 10.1016/j.phytochem.2007.07.021
|
| [29] | Sarg T, El-Dahmy SI, Salem S (1990) Germacranolides from Anthemis melampodina. Sci Pharm 58: 33-35. |
| [30] |
Ghantous A, Sinjab A, Herceg Z, et al. (2013) Parthenolide: from plant shoots to cancer roots. Drug Discov Today 18: 894-905. doi: 10.1016/j.drudis.2013.05.005
|
| [31] |
Yang Q, Wan L, Zhou Z, et al. (2013) Parthenolide from Parthenium integrifolium reduces tumor burden and alleviate cachexia symptoms in the murine CT-26 model of colorectal carcinoma. Phytomedicine 20: 992-998. doi: 10.1016/j.phymed.2013.04.020
|
| [32] | Wollenweber E, Schober I, Dostal P, et al. (1986) Flavonoids and terpenoids from the exudates of some Baccharis species. Zeitschrift fuer Naturforschung C 41: 87-93. |
| [33] | Le Quesne PW, Levery SB, Menachery MD, et al. (1979) ChemInform Abstract: Antitumor plants Part 6. Novel modified germacranolides and other constituents of Eremanthus elaeagnus Schultz-Bip (Compositae). ChemInform 12: 1572-1580. |
| [34] | Saito Y, Iwamoto Y, Okamoto Y, et al. (2012) Four new guaianolides and acetylenic alcohol from Saussurea katochaete collected in China. Nat Prod Commun 7: 447-450. |
| [35] | Ahmed KM, El-Din SS, Wahab SA, et al. (2001) Study of the coumarin and volatile oil composition from aerial parts of Achillea millefolium L. Pak J Sci Ind Res 44: 218-222. |
| [36] | Gonzalez AG, Estevez RR, Herrera VJ (1975) Chemistry of the Compositae. XXVI. Sesquiterpene lactones and coumarins of Artemisia ramosa. Anales de Quimica 71: 437-439. |
| [37] | Hsieh SF, Hsieh TJ, El-Shazly M, et al. (2012) Chemical constituents from Farfugium japonicum var. formosanum. Nat Prod Commun 7: 435-440. |
| [38] | Li YS, Luo SD, Zhang M, et al. (2001) Constituents of Liguliria vellerea (Franch.) Hand-Mazz. China J Chin Mater Med 26: 835-837. |
| [39] | Plouvier V (1963) Distribution of aliphatic polyols and cyclitols. In Chemical Plant Taxonomy, 1st Edition, Swain T. Ed, Academic Press London - New York. Chap. 11, pp 321. |
| [40] |
Abe F, Yamauchi T, Honda K, et al. (1998) Cyclitols and their glycosides from leaves of Marsdenia tomentosa. Phytochemistry 47: 1297-1301. doi: 10.1016/S0031-9422(97)00766-8
|
| [41] |
Merchant A, Richter A, Popp M, et al. (2006) Targeted metabolite profiling provides a functional link among eucalypt taxonomy, physiology and evolution. Phytochemistry 67: 402-408. doi: 10.1016/j.phytochem.2005.11.027
|
| [42] |
Adams MA, Richter A, Hill AK, et al. (2005) Salt tolerance in Eucalyptus spp.: identity and response of putative osmolytes. Plant Cell Environ 28: 772-787. doi: 10.1111/j.1365-3040.2005.01328.x
|
| [43] |
Honda K, Omura H, Hayashi N, et al. (2004) Conduritols as oviposition stimulants for the danaid butterfly, Parantica sita, identified from a host plant, Marsdenia tomentosa. J Chem Ecol 30: 2285-2296. doi: 10.1023/B:JOEC.0000048789.06504.f1
|
| [44] | Kim SL, Lee ST, Trang KT, et al. (2014) Parthenolide exerts inhibitory effects on angiogenesis through the downregulation of VEGF/VEGFRs in colorectal cancer. Int J Mol Med 33: 1261-1267. |
| [45] |
Al-Fatlawi AA, Al-Fatlawi AA, Irshad M, et al. (2015) Effect of parthenolide on growth and apoptosis regulatory genes of human cancer cell lines. Pharm Biol 53:104-109. doi: 10.3109/13880209.2014.911919
|
| [46] | Dong L, Qiao H, Zhang X, et al. (2013) Parthenolide is neuroprotective in rat experimental stroke model: downregulating NF-κB, phospho-p38MAPK, and caspase-1 and ameliorating BBB permeability. Mediat Inflamm 2013: 370804. |
| [47] |
Materazzi S, Benemei S, Fusi C, et al. (2013) Parthenolide inhibits nociception and neurogenic vasodilatation in the trigeminovascular system by targeting the TRPA1 channel. Pain 154: 2750-2578. doi: 10.1016/j.pain.2013.08.002
|
| [48] |
Magni P, Ruscica M, Dozio E, et al. (2012) Parthenolide inhibits the LPS-induced secretion of IL-6 and TNF-α and NF-κB nuclear translocation in BV-2 microglia. Phytother Res 26: 1405-1409. doi: 10.1002/ptr.3732
|
| [49] | Nicolini F, Burmistrova O, Marrero MT, et al. (2014) Induction of G2/M phase arrest and apoptosis by the flavonoid tamarixetin on human leukemia cells. Mol Carcinog 53: 939-950. |
| [50] |
Ezenyi IC, Salawu OA, Kulkarni R, et al. (2014) Antiplasmodial activity-aided isolation and identification of quercetin-4'-methyl ether in Chromolaena odorata leaf fraction with high activity against chloroquine-resistant Plasmodium falciparum. Parasitol Res 113: 4415-4422. doi: 10.1007/s00436-014-4119-y
|
| [51] |
Lau AJ, Chang TK (2013) Indirect activation of the SV23 and SV24 splice variants of human constitutive androstane receptor: analysis with 3-hydroxyflavone and its analogues. Br J Pharmacol 170: 403-414. doi: 10.1111/bph.12284
|
| [52] |
Lau AJ, Chang TK (2015) 3-Hydroxyflavone and structural analogues differentially activate pregnane X receptor: Implication for inflammatory bowel disease. Pharmacol Res 100: 64-72. doi: 10.1016/j.phrs.2015.07.031
|
| [53] | Al-Majedy YK, Al-Duhaidahawi DL, Al-Azawi KF, et al. (2016) Coumarins as Potential Antioxidant Agents Complemented with Suggested Mechanisms and Approved by Molecular Modeling Studies. Molecules 21: E135. |
| [54] |
Jagadeesh GS, Nagoor Meeran MF, Selvaraj P (2016) Protective Effects of 7-Hydroxycoumarin on Dyslipidemia and Cardiac Hypertrophy in Isoproterenol-Induced Myocardial Infarction in Rats. J Biochem Mol Toxicol 30: 120-127. doi: 10.1002/jbt.21770
|
| [55] | Soto-Nuñez M, Díaz-Morales KA, Cuautle-Rodríguez P, et al. (2015) Single-cell microinjection assay indicates that 7-hydroxycoumarin induces rapid activation of caspase-3 in A549 cancer cells. Exp Ther Med 10: 1789-1795. |
| [56] |
Alvarez MA, Rotelli AE, Pelzer LE, et al. (2000) Phytochemical study and anti-inflammatory properties of Lampaya hieronymi Schum. Ex Moldenke. Il Farmaco 55: 502-505. doi: 10.1016/S0014-827X(00)00067-7
|
| [57] |
Billington DC, Perron-Sierra F, Picard I, et al. (1994) Total synthesis of novel conduritol related compounds capable of modulating insulin release. Bioorg Med Chem Lett 4: 2307-2312. doi: 10.1016/0960-894X(94)85030-5
|
Figures(1)

Alessandro Venditti, Claudio Frezza, Giacomo Rossi, Mirella Di Cecco, Giampiero Ciaschetti, Mauro Serafini, Armandodoriano Bianco. Secondary metabolites with ecologic and medicinal implications in Anthemis cretica subsp. petraea from Majella National Park[J]. AIMS Molecular Science, 2016, 3(4): 648-660. doi: 10.3934/molsci.2016.4.648

 DownLoad:
DownLoad:
