Citation: Masaki Sekiguchi, Emiko Ishiwata, Yukihiko Nakata. Dynamics of an ultra-discrete SIR epidemic model with time delay[J]. Mathematical Biosciences and Engineering, 2018, 15(3): 653-666. doi: 10.3934/mbe.2018029

| [1] | [ L. J. S. Allen, Some discrete-time SI, SIR and SIS epidemic models, Math. Bio., 124 (1994): 83-105. |
| [2] | [ E. Beretta,Y. Takeuchi, Global stability of an SIR epidemic model with time delays, J. Math. Biol., 33 (1995): 250-260. |
| [3] | [ E. Beretta,T. Hara,W. Ma,Y. Takeuchi, Global asymptotic stability of an SIR epidemic model with distributed time delay, Nonl. Anal., 47 (2001): 4107-4115. |
| [4] | [ R. M. Corless,C. Essex,M. A. H. Nerenberg, Numerical methods can suppress chaos, Phys. Lett. A, 157 (1991): 27-36. |
| [5] | [ Y. Enatsu,Y. Nakata,Y. Muroya, Global stability for a class of discrete SIR epidemic models, Math. Bio. and Eng., 7 (2010): 347-361. |
| [6] | [ Y. Enatsu,Y. Nakata,Y. Muroya,G Izzo,A Vecchio, Global dynamics of difference equations for SIR epidemic models with a class of nonlinear incidence rates, J. Diff. Equ. Appl., 18 (2012): 1163-1181. |
| [7] | [ S. Elaydi, An Introduction to Difference Equations, Springer-Verlag, New York, 2005. |
| [8] | [ D. F. Griffiths,P. K. Sweby,H. C. Yee, On spurious asymptotic numerical solutions of explicit Runge-Kutta methods, IMA J. Numer. Anal., 12 (1992): 319-338. |
| [9] | [ J. K. Hale and S. M. Verduyn Lunel, Introduction to Functional Differential Equations, Applied Mathematical Sciences Vol 99, Springer, 1993. |
| [10] | [ G. Izzo,A. Vecchio, A discrete time version for models of population dynamics in the presence of an infection, J. Comput. Appl. Math., 210 (2007): 210-221. |
| [11] | [ G. Izzo, Y. Muroya and A. Vecchio, A general discrete time model of population dynamics in the presence of an infection Disc. Dyn. Nat. Soc. , (2009), Art. ID 143019, 15pp. |
| [12] | [ L. Jódar,R. J. Villanueva,A. J. Arenas,G. C. González, Nonstandard numerical methods for a mathematical model for influenza disease, Math. Comput. Simul., 79 (2008): 622-633. |
| [13] | [ C. M. Kent, Piecewise-defined difference equations: Open Problem, 'Bridging Mathematics, Statistics, Engineering and Technology, 55-71, Springer Proc. Math. Stat., 24, Springer, New York, 2012. |
| [14] | [ C. C. McCluskey, Complete global stability for an SIR epidemic model with delay -distributed or discrete time delays, Nonl. Anal. RWA, 11 (2010): 55-59. |
| [15] | [ R. E. Mickens, Discretizations of nonlinear differential equations using explicit nonstandard methods, J. Comput. Appl. Math., 110 (1999): 181-185. |
| [16] | [ S. M. Moghadas,M. E. Alexander,B. D. Corbett,A. B. Gumel, A positivity-preserving Mickens-type discretization of an epidemic model, J. Diff. Equ. Appl., 9 (2003): 1037-1051. |
| [17] | [ K. Matsuya,M. Murata, Spatial pattern of discrete and ultradiscrete Gray-Scott model, DCDS-B, 20 (2015): 173-187. |
| [18] | [ K. Matsuya and M. Kanai, Exact solution of a delay difference equation modeling traffic flow and their ultra-discrete limit, arXiv: 1509.07861 [nlin. CG]. |
| [19] | [ K. Nishinari,D. Takahashi, Analytical properties of ultradiscrete Burgers equation and rule-184 cellular automaton, J. Phys. A, 31 (1998): 5439-5450. |
| [20] | [ A. Ramani,A. S. Carstea,R. Willox,B. Grammaticos, Oscillating epidemics: A discrete-time model, Phys. A, 333 (2004): 278-292. |
| [21] | [ T. Tokihiro,D. Takahashi,J. Matsukidaira,J. Satsuma, From soliton equations to integrable cellular automata through a limiting procedure, Phys. Rev. Lett., 76 (1996): 3247-3250. |
| [22] | [ J. Satsuma,R. Willox,A. Ramani,B. Grammaticos,A. S. Carstea, Extending the SIR epidemic model, Phys. A, 336 (2004): 369-375. |
| [23] | [ M. Sekiguchi, Permanence of some discrete epidemic models, Int. J. Biomath., 2 (2009): 443-461. |
| [24] | [ M. Sekiguchi,E. Ishiwata, Global dynamics of a discretized SIRS epidemic model with time delay, J. Math. Anal. Appl., 371 (2010): 195-202. |
| [25] | [ G. C. Sirakoulis,I. Karafyllidis,A. Thanailakis, A cellular automaton model for the effects of population movement and vaccination on epidemic propagation, Ecol. Mod., 133 (2000): 209-223. |
| [26] | [ S. H. White,A. Martin del Rey,G. Rodríguez Sánchez, Modeling epidemics using cellular automata, Appl. Math. Comp., 186 (2007): 193-202. |
| [27] | [ R. Willox,B. Grammaticos,A. S. Carstea,A. Ramani, Epidemic dynamics: Discrete-time and cellular automaton models, Phys. A, 328 (2003): 13-22. |
| [28] | [ S. Wolfram, Statistical mechanics of cellular automata, Rev. Mod. Phys., 55 (1983): 601-644. |
| [29] | [ T. Zhang,Z. Teng, Global behavior and permanence of SIRS epidemic model with time delay, Nonl. Anal. RWA., 9 (2008): 1409-1424. |
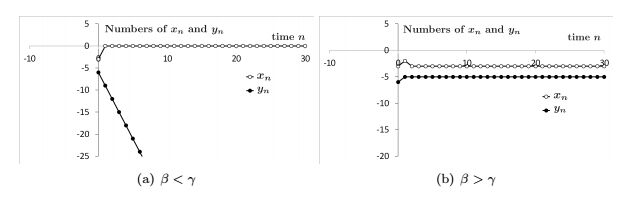
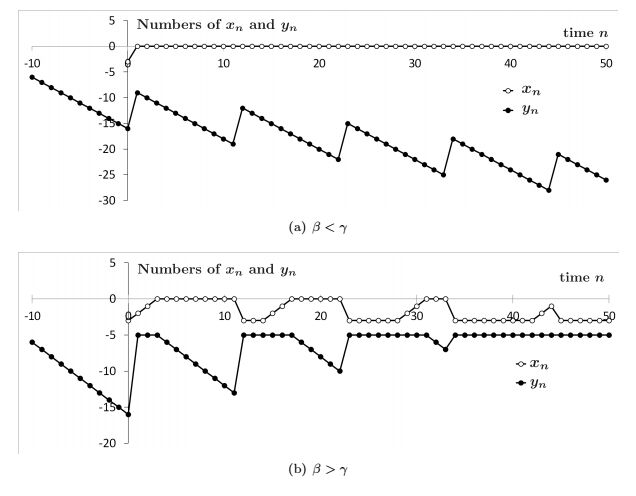
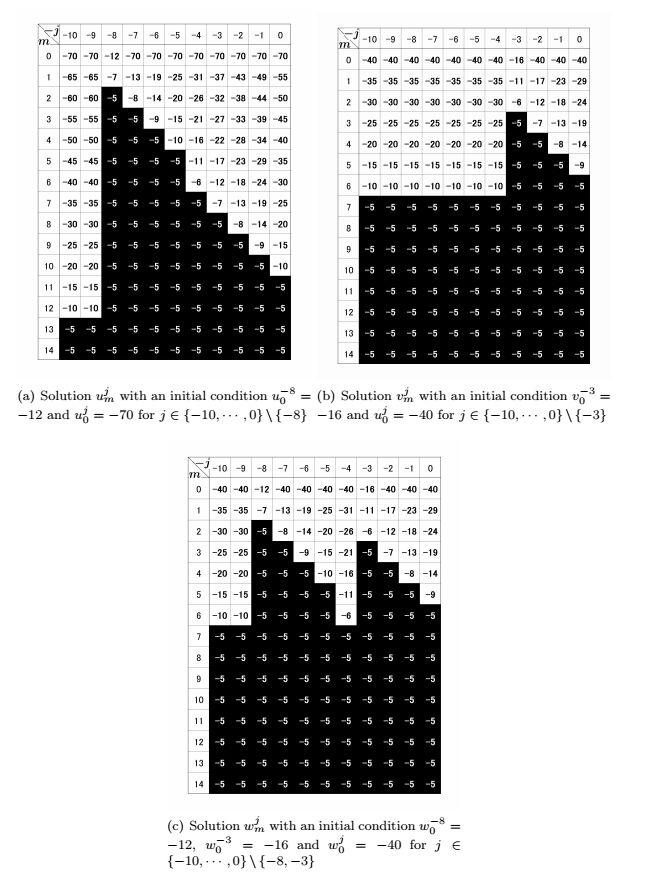
Figures(3)

Masaki Sekiguchi, Emiko Ishiwata, Yukihiko Nakata. Dynamics of an ultra-discrete SIR epidemic model with time delay[J]. Mathematical Biosciences and Engineering, 2018, 15(3): 653-666. doi: 10.3934/mbe.2018029







 DownLoad:
DownLoad: