Citation: Moven Manjowe, Terence Darlington Mushore, Juliet Gwenzi, Collen Mutasa, Electdom Matandirotya, Emmanuel Mashonjowa. Circulation mechanisms responsible for wet or dry summers over Zimbabwe[J]. AIMS Environmental Science, 2018, 5(3): 154-172. doi: 10.3934/environsci.2018.3.154

| [1] |
ADDIN Mendeley Bibliography CSL_BIBLIOGRAPHY Moyo EN, Nangombe SS (2015) Southern Africa's 2012–13 violent storms: role of climate change. Procedia IUTAM 17: 69–78. doi: 10.1016/j.piutam.2015.06.011

|
| [2] | Wahid H, Ahmad S, Nor MSM, et al. (2017) Prestasi kecekapan pengurusan kewangan dan agihan zakat: perbandingan antara majlis agama islam negeri di Malaysia. J Ekon Malaysia 51: 39–54. |
| [3] | Mutasa M (2010) Zimbabwe' s Drought Conundrum : vulnerability and coping in Buhera and Chikomba districts. MS thesis, 163. |
| [4] | Ndlovu S (2010) Coping With Drought. Mainstreaming Livelihood Centred Approaches to Disaster Management. |
| [5] |
Levey KM, Jury MR (1996) Composite intraseasonal oscillations of convection over Southern Africa. J Clim 9: 1910–1920. doi: 10.1175/1520-0442(1996)009<1910:CIOOCO>2.0.CO;2
|
| [6] | Makarau A (1995) Intra-seasonal oscillatory modes of the southern Africa summer circulation, Univ. Cape T. (PhD thesis). |
| [7] | Zhakata W (1996) Impacts of climate variability and forecasting on agriculture (Zimbabwe experience), in Proc. Workshop on Reducing Climate-Related Vulnerability in Southern Africa, 131–139. |
| [8] | Jury MR (1993) The structure and possible forcing mechanisms of the 1991–1992 drought in Southern Africa, Suid-Afrikaanse Tydskr. vir Natuurwetenskap en Tegnol 12, 8–16. |
| [9] |
Colberg F (2004) South Atlantic response to El Niño–Southern Oscillation induced climate variability in an ocean general circulation model. J Geophys Res 109: C12015. doi: 10.1029/2004JC002301
|
| [10] |
Chikozho C (2010) Applied social research and action priorities for adaptation to climate change and rainfall variability in the rainfed agricultural sector of Zimbabwe. Phys Chem Earth 35: 780–790. doi: 10.1016/j.pce.2010.07.006
|
| [11] | Gwimbi P (2009) Cotton farmers' vulnerability to climate change in Gokwe District (Zimbabwe): impact and influencing factors. JÀMBÁ J Disaster Risk Stud 2: 81–92. |
| [12] | Alvarez TC, Delgado RC, González PA (2014) Aplicabilidad de los sistemas de triaje prehospitalarios en los incidentes con múltiples víctimas: De la teoría a la práctica. Emergencias 26: 147–154. |
| [13] | Tome S (2017) Weekly Bulletin on Outbreaks. Week 46, no. July, 1–7. |
| [14] | Bohle HG, Downing TE, Watts M J (1994) Climate Change and Social Vulnerability. Toward a Sociology and Geography of Food Insecurity. Glob Environ Chang 4: 37–48. |
| [15] |
Mavhura E, Bernard S, Collins AE, et al. (2013) Author' s personal copy International Journal of Disaster Risk Reduction Indigenous knowledge, coping strategies and resilience to floods in Muzarabani, Zimbabwe. Int J Disaster Risk Reduct 5: 38–48. doi: 10.1016/j.ijdrr.2013.07.001
|
| [16] | Mavhura E, Manatsa D, Mushore T (2015) Adaptation to drought in arid and semi-arid environments: Case of the Zambezi Valley, Zimbabwe. Jàmbá J Disaster Risk Stud 7: 1–7. |
| [17] |
Mushore TD (2013) Uptake Of Seasonal Rainfall Forecasts In Zimbabwe. IOSR J Environ Sci Toxicol Food Technol 5: 31–37. doi: 10.9790/2402-0513137
|
| [18] |
Cook KH (2000) The South Indian convergence zone and interannual rainfall variability over Southern Africa. J Clim 13: 3789–3804. doi: 10.1175/1520-0442(2000)013<3789:TSICZA>2.0.CO;2
|
| [19] | Mason S, Lindesay J, Tyson J (1994) Simulating drought in Southern Africa using sae surface temperature variations. Water SA 20: 15–22. |
| [20] | Beilfuss R (2012) A Risky Climate for Southern African Hydro. Int Rivers Berkely, 1–46. |
| [21] |
Mamombe V, Kim WM, Choi YS (2017) Rainfall variability over Zimbabwe and its relation to large-scale atmosphere–ocean processes. Int J Climatol 37: 963–971. doi: 10.1002/joc.4752
|
| [22] | Mwafulirwa ND (1999) Climate variability and predictability in tropical southern africa with a focus on dry spells over Malawi. |
| [23] |
Manatsa D, Chingombe W, Matsikwa H, et al. (2008) The superior influence of Darwin Sea level pressure anomalies over ENSO as a simple drought predictor for Southern Africa. Theor Appl Climatol 92: 1–14. doi: 10.1007/s00704-007-0315-3
|
| [24] | Manatsa D, Mukwada G (2012) Rainfall Mechanisms for the Dominant Rainfall Mode over Zimbabwe Relative to ENSO and/or IODZM. Sci World J 2012: 1–15. |
| [25] |
Gaughan AE, Staub CG, Hoell A, et al. (2016) Inter- and Intra-annual precipitation variability and associated relationships to ENSO and the IOD in southern Africa. Int J Climatol 36: 1643–1656. doi: 10.1002/joc.4448
|
| [26] | Ogwang BA, Guirong T, Haishan C (2012) Diagnosis of September - November Drought and the Associated Circulation Anomalies Over Uganda. Pakistan J Meteorol 9: 11–24. |
| [27] |
Kabanda TA, Jury MR (1999) Inter-annual variability of short rains over northern Tanzania. Clim Res 13: 231–241. doi: 10.3354/cr013231
|
| [28] |
Nangombe S, Madyiwa S, Wang J (2018) Precursor conditions related to Zimbabwe's summer droughts. Theor Appl Climatol 131: 413–431. doi: 10.1007/s00704-016-1986-4
|
| [29] |
Manatsa D, Reason CJC, Mukwada G (2012) On the decoupling of the IODZM from southern Africa Summer rainfall variability. Int J Climatol 32: 727–746. doi: 10.1002/joc.2306
|
| [30] |
Simmonds I, Hope P (1997) Persistence Characteristics of Australian Rainfall Anomalies. Int J Climatol 17: 597–613. doi: 10.1002/(SICI)1097-0088(199705)17:6<597::AID-JOC173>3.0.CO;2-V
|
| [31] |
Manatsa D, Mushore T, Lenouo A (2017) Improved predictability of droughts over southern Africa using the standardized precipitation evapotranspiration index and ENSO. Theor Appl Climatol 127: 259–274. doi: 10.1007/s00704-015-1632-6
|
| [32] | Department of Meterological Services (1981) Climate Handbook of Zimbabwe, Zimbabwe Meteorol. Serv. Harare, 48. |
| [33] |
Mavhura E, Collins A, Bongo PP (2017) Flood vulnerability and relocation readiness in Zimbabwe. Disaster Prev Manag An Int J 26: 41–54. doi: 10.1108/DPM-05-2016-0101
|
| [34] | Muzenda-Mudavanhu C, Manyena B, Collins AE (2015) Disaster risk reduction knowledge among children in Muzarabani District, Zimbabwe. Nat Hazards 84: 911–931. |
| [35] | Bohle HG, Downing TE, Watts MJ (1994) Köppen Climate Classification for Bucharest. |
| [36] | Janus Corporate Solutions (2016) Weather and Climate in Singapore. Longman. |
| [37] | Matarira CH (1990) Theoretical and Ap. plied Climatology Frequency and Tracks of Anticyclones and Their Effect on Rainfall Pat- terns over Zimbabwe. Theor Appl Climatol 66: 53–66. |
| [38] |
Makarau A, Jury MR (1997) Seasonal Cycle of Convective Spells Over Southern. Int J Climatol 17: 1317–1332. doi: 10.1002/(SICI)1097-0088(199710)17:12<1317::AID-JOC197>3.0.CO;2-A
|
| [39] |
Manatsa D, Morioka Y, Behera SK, et al. (2013) Link between Antarctic ozone depletion and summer warming over southern Africa. Nat Geosci 6: 934–939. doi: 10.1038/ngeo1968
|
| [40] | Levitus S, Boyer T (1998) NOAA/OAR/ESRL PSD, Boulder, Colorado, USA. |
| [41] |
Dee DP, Uppala SM, Simmons AJ, et al. (2011) The ERA-Interim reanalysis: Configuration and performance of the data assimilation system. Q J R Meteorol Soc 137: 553–597. doi: 10.1002/qj.828
|
| [42] |
Kalnay E, Kanamitsu M, Kistler R, et al. (1996) The NCEP/NCAR 40-year reanalysis project. Bull Am Meteorol Soc 77: 437–471. doi: 10.1175/1520-0477(1996)077<0437:TNYRP>2.0.CO;2
|
| [43] |
Smith TM, Reynolds RW, Peterson TC, et al. (2008) Improvements to NOAA's historical merged land–ocean surface temperature analysis (1880–2006). J Clim 21: 2283–2296. doi: 10.1175/2007JCLI2100.1
|
| [44] | Folland CK (1983) Regional-Scale International Variability Of Climate-A Northwest European-Perspective. Meteorol Mag 112: 163–183. |
| [45] | Mpeta EJ (1997) Intra-seasonal convection dynamics over southwest and northeast Tanzania: an observational study, University of Cape Town. |
| [46] | Naeraa M, Jury MR (1998) Meteorology and Atmospheric Physics Tropical Cyclone Composite Structure and Impacts over Eastern Madagascar During January-March 1994. Meteorol Atmos Phys 53: 43–53. |
| [47] | Jury MR, Levey KM (1997) Vertical structure of the atmosphere during wet spells over Southern Africa. Water SA 23: 51–55. |
| [48] | Hargraves R, Jury M (1997) Composite meteorological structure of flood events over the eastern mountains of South Africa. Water SA 23: 357–363. |
| [49] |
Boschat G, Simmonds L, Purich A, et al. (2016) On the use of composite analyses to form physical hypotheses: An example from heat wave-SST associations. Sci Rep 6: 29599. doi: 10.1038/srep29599
|
| [50] | Rocha A, Simmonds I (1997) Interannual Variability of South-Eastern African Summer Rainfall. Part Ii. Modelling the Impact of Sea-Surface Temperatures on Rainfall and Circulation. Int J Climatol 17: 267–290. |
| [51] |
Simmonds I, Bi D, Hope P (1999) Atmospheric water vapor flux and its association with rainfall over China in summer. J Clim 12: 1353–1367. doi: 10.1175/1520-0442(1999)012<1353:AWVFAI>2.0.CO;2
|
| [52] | Preston-Whyte RA, Tyson PD (1988) The atmosphere and weather of Southern Africa. Oxford University Press. |
| [53] | Rocha A, Simmonds I (1997) Interannual Variability of South-Eastern African Summer Rainfall. Part 1: Relationships With Air–Sea Interaction Processes. Int J Climatol 17: 235–265. |
| [54] |
Nicholson SE, Entekhabi D (1987) Rainfall Variability in Equatorial and Southern Africa: Relationships with Sea Surface Temperatures along the Southwestern Coast of Africa. J Clim Appl Meteorol 26: 561–578. doi: 10.1175/1520-0450(1987)026<0561:RVIEAS>2.0.CO;2
|
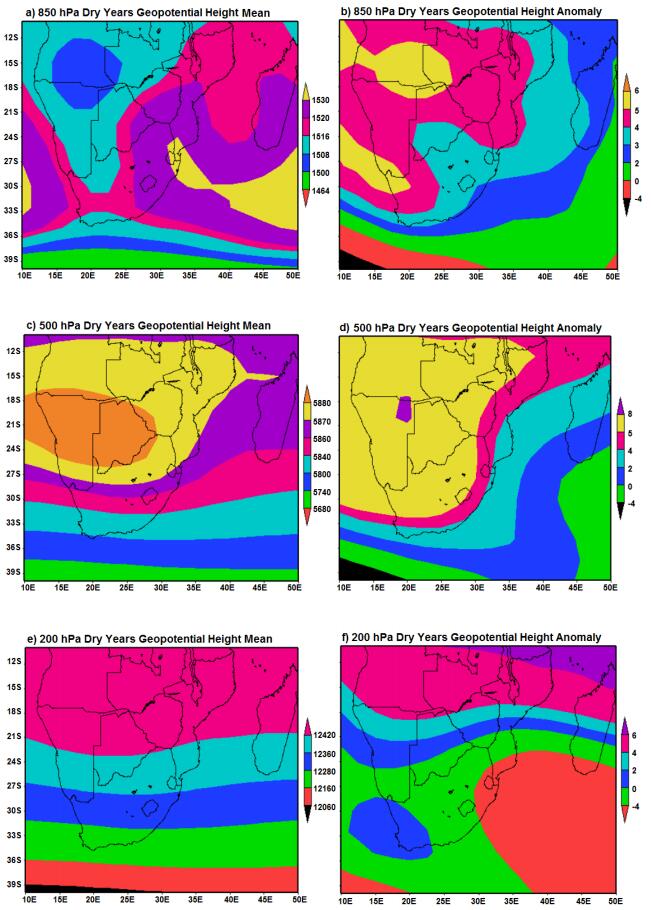
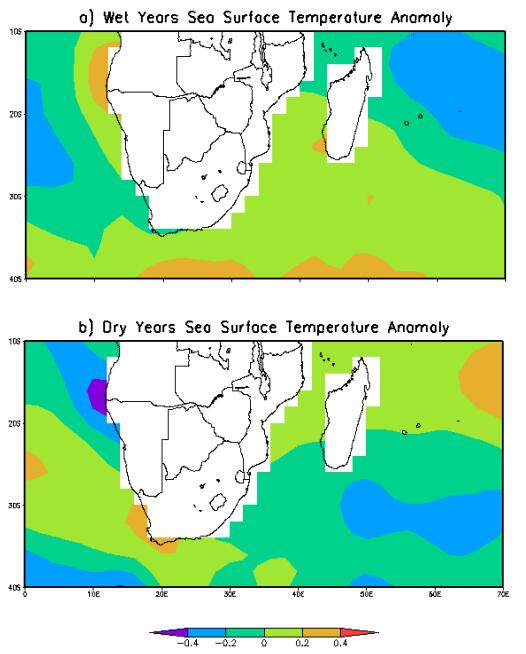
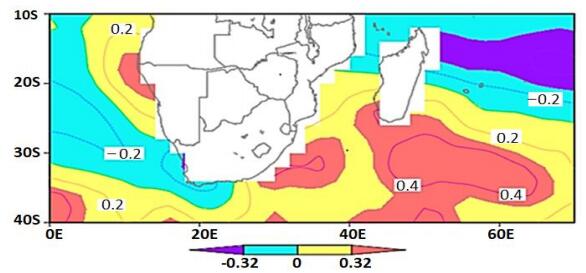
Figures(7)

Moven Manjowe, Terence Darlington Mushore, Juliet Gwenzi, Collen Mutasa, Electdom Matandirotya, Emmanuel Mashonjowa. Circulation mechanisms responsible for wet or dry summers over Zimbabwe[J]. AIMS Environmental Science, 2018, 5(3): 154-172. doi: 10.3934/environsci.2018.3.154







 DownLoad:
DownLoad: