Citation: Joanna Faber, Krzysztof Brodzik. Air quality inside passenger cars[J]. AIMS Environmental Science, 2017, 4(1): 112-133. doi: 10.3934/environsci.2017.1.112

| [1] | Glader A (2012) Chemical emissions from building structures – Emission sources and their impact on indoor air. Department of Public Health and Clinical Medicine, Umeå, Sweden. |
| [2] | Levin H (2003) Indoor air pollutants. Part 1: general description of pollutants, levels and standards. Ventilation Information Paper 2. |
| [3] | Minister of Supply and Services, Canada (1995) Exposure guidelines for residential indoor air quality. A Report of the Federal-Provincial Advisory Committee on Environmental and Occupational Health. |
| [4] | Report No 19, EUR 17675 EN (1997) Total volatile organic compounds (TVOC) in indoor air quality investigations. European Collaborative Action 'Indoor Air Quality & its Impact on Man', Luxembourg. |
| [5] | ISO 16000-6:2011: Determination of volatile organic compounds in indoor and test chamber air by active sampling on Tenax TA® sorbent, thermal desorption and gas chromatography using MS or MS-FID. |
| [6] |
Jones AP (1999) Indoor air quality and health. Atmos Environ 33: 4535-4564. doi: 10.1016/S1352-2310(99)00272-1

|
| [7] |
Weschler ChJ, Nazaroff WW (2008) Semivolatile organic compounds in indoor environments. Atmos Environ 42: 9018-9040. doi: 10.1016/j.atmosenv.2008.09.052
|
| [8] | Heinzow B, Sagunski H (2009) Evaluation of indoor air contamination by means of reference and guide values: the German approach, In: Salthammer T, Uhde E, Organic indoor air pollutants 2 ed. Weinheim: Wiley-VCH Verlag GmbH, 189-211. |
| [9] | Sărbu I, Sebarchievici C (2011) Olfactory comfort assurance in buildings, In: Mazzeo N (ed.), Chemistry, emission control, radioactive pollution and indoor air quality, InTech, 407-428. |
| [10] | Dinh TV, Choi IY, Son YS, et al. (2016) Volatile organic compounds (VOCs) in surface coating materials: their compositions and potential as an alternative fuel. J Environ Manage 168: 157-164. |
| [11] | Selected pollutants (2010) WHO Guidelines for Indoor Air Quality, WHO. |
| [12] |
Schupp T, Bolt HM, Jaeckh R, et al. (2006) Benzene and its methyl-derivatives: Derivation of maximum exposure levels in automobiles. Toxicol Lett 160: 93-104. doi: 10.1016/j.toxlet.2005.06.012
|
| [13] | Wadge A, Salisbury J (1997) Benzene. National Environmental Health Monographs. Air Series No. 2, Department of Human Services, Australia, Glenelg Press. |
| [14] | World Health Organization (1999) Indoor air quality: organic pollutants. EURO Reports and Studies 111, Copenhagen, Denmark. |
| [15] | Mapou AEM, Shendell DG, Therkorn JH (2006) Aldehydes in passenger vehicles: An analysis of data from the RIOPA study 1999-2001. Atmos Environ 79: 751-759. |
| [16] | Air Quality Sciences, Inc. (2006) Indoor air quality hazards of new cars. Marietta, USA. |
| [17] | Xu B, Wu Y, Gong Y, et al. (2016) Investigation of volatile organic compounds exposure inside vehicle cabins in China. Atmos Pollut Res 7: 215-220. |
| [18] |
Kalman EL, Löfvendahl A, Winquist F, et al. (2000) Classification of complex gas mixtures from automotive leather using an electronic nose. Anal Chim Acta 403: 31-38. doi: 10.1016/S0003-2670(99)00604-2
|
| [19] |
Wensing M, Uhde E, Salthammer T (2005) Plastic additives in the indoor environment – flame retardants and plasticizers. Sci Total Environ 339: 19-40. doi: 10.1016/j.scitotenv.2004.10.028
|
| [20] | Johansson I (1999) The role of volatile organic compounds in the assessment of indoor air quality. Karolinska Institutet, Stockholm, Sweden. |
| [21] |
Delahunty CM, Eyres G, Dufour JP (2006) Gas chromatography-olfactometry. J Separ Sci 29: 2107-2125. doi: 10.1002/jssc.200500509
|
| [22] |
Bartsch J, Uhde E, Salthammer T (2016) Analysis of odour compounds from scented consumer products using gas chromatography-mass spectrometry and gas chromatography-olfactometry. Anal Chim Acta 904: 98-106. doi: 10.1016/j.aca.2015.11.031
|
| [23] |
Mølhave L, Clausen G, Berglund B, et al. (1997) Total volatile organic compounds (TVOC) in indoor air quality investigations. Indoor Air 7: 225-240. doi: 10.1111/j.1600-0668.1997.00002.x
|
| [24] |
Mandalakis M, Stephanou EG, Horii Y, et al. (2008) Emerging contaminants in car interiors: evaluating the impact of airborne PBDEs and PBDD/Fs. Environ Sci Technol 42: 6431-6436. doi: 10.1021/es7030533
|
| [25] | Yoshida T, Matsunaga I, Tomioka K, et al. (2006) Interior air pollution in automotive cabins by volatile organic compounds diffusing from interior materials: I. Survey of 101 types of Japanese domestically produced cars for private use. Indoor Built Environ 15: 425-444. |
| [26] |
Jo WK, Yu CH (2001) Public bus and taxicab drivers' exposure to aromatic work-time Volatile Organic Compounds. Environ Res A 86: 66-72. doi: 10.1006/enrs.2001.4257
|
| [27] |
Batterman SA, Peng ChY, Braun J (2002) Levels and composition of volatile organic compounds on commuting routes in Detroit, Michigan. Atmos Environ 36: 6015-6030. doi: 10.1016/S1352-2310(02)00770-7
|
| [28] |
Jo WK, Choi SJ (1996) Vehicle occupants' exposure to aromatic Volatile Organic Compounds while commuting on an urban-suburban route in Korea. J Air Waste Manage Assoc 46: 749-754. doi: 10.1080/10473289.1996.10467509
|
| [29] | Weisel CP (2005) Automobile, bus and rail passenger air quality. In: Martin Hocking, editor, The Handbook of Environmental Chemistry, Springer Berlin Heidelberg, 4H: 317-334. |
| [30] |
Jo WK, Park KH (1998) Concentrations of volatile organic compounds in automobile's cabins while commuting along a Korean urban area. Environ Int 24: 259-265. doi: 10.1016/S0160-4120(98)00004-X
|
| [31] |
Riediker M, Williams R, Devlin R, et al. (2003) Exposure to particulate matter, volatile organic compounds and other air pollutants inside patrol cars. Environ Sci Technol 37: 2084-2093. doi: 10.1021/es026264y
|
| [32] |
Jo WK, Park KH (1999) Concentrations of volatile organic compounds in the passenger side and the back seat of automobiles. J Expo Anal Environ Epid 9: 217-227. doi: 10.1038/sj.jea.7500041
|
| [33] | Grabbs JS, Corsi RL, Torres VM (2000) Volatile organic compounds in new automobiles: screening assessment. J Environ Eng 974-977. |
| [34] |
Schlink U, Rehwagen M, Damm M, et al. (2004) Seasonal cycle of indoor-VOCs: comparison of apartments and cities. Atmos Environ 38: 1181-1190. doi: 10.1016/j.atmosenv.2003.11.003
|
| [35] |
Yoshida T, Matsunaga I (2006) A case study on identification of airborne organic compounds and time courses of their concentration in the cabin of a new car for private use. Environ Int 32: 58-79. doi: 10.1016/j.envint.2005.04.009
|
| [36] | Ilgen E, Levsen K, Angerer J, et al. (2001) Aromatic hydrocarbons in the atmospheric environment. Part III: personal monitoring. Atmos Environ 35: 1265-1279. |
| [37] |
Chien YCh (2007) Variations in amounts and potential sources of volatile organic chemicals in new cars. Sci Total Environ 382: 228-239. doi: 10.1016/j.scitotenv.2007.04.022
|
| [38] |
You KW, Ge YS, Hu B, et al. (2007) Measurement of in-vehicle volatile organic compounds under static conditions. J Environ Sci 19: 1208-1213. doi: 10.1016/S1001-0742(07)60197-1
|
| [39] | Cho Y, Park DS, Kwon SB (2015) Characterization of VOC emissions from interior materials of railroad passenger cabin and preparation of environment-friendly interior materials. Korea Railroad Research Institute. |
| [40] |
Fedoruk MJ, Kerger BD (2003) Measurement of volatile organic compounds inside automobiles. J Expo Anal Environ Epid 13: 31-41. doi: 10.1038/sj.jea.7500250
|
| [41] | Fung W, Hardcastle M (2001) Textiles in automotive engineering. The Textile Institute, Cambridge, England: Woodhead Publishing Ltd. |
| [42] | Tucker WG (2001) Volatile organic compounds, In: Spengler JD, Samet JM, McCarthy JF, editors, Indoor Air Quality Handbook, The McGraw – Hill Companies Inc., 31.1-31.20. |
| [43] |
Yeganeh B, Haghighat F, Gunnarsen L, et al. (2006) Evaluation of building materials individually and in combination using odour threshold. Indoor Built Environ 15: 583-593. doi: 10.1177/1420326X06072735
|
| [44] |
Setnikova I (2014) Impact of indoor surface material on perceived air quality. Mat Sci Eng C 36: 1-6. doi: 10.1016/j.msec.2013.11.032
|
| [45] | Kataoka H, Ohashi Y, Mamiya T, et al. (2012) Indoor air monitoring of volatile organic compounds and evaluation of their emission from various building materials and common products by gas chromatography-mass spectrometry. Advanced Gas Chromatography – Progress in Agricultural, Biomedical and Industrial Applications. InTech. |
| [46] |
Wolkoff P (1999) How to measure and evaluate volatile organic compound emissions from building products. A perspective. Sci Total Environ 227: 197-213. doi: 10.1016/S0048-9697(99)00019-4
|
| [47] | Zabiegała B (2006) Organic compounds in indoor environments. Pol J Environ Stud 15: 383-393. |
| [48] |
Lin CC, Yu KP, Zhao P, et al. (2009) Evaluation of impact factors on VOC emissions and concentrations from wooden flooring based on chamber tests. Build Environ 44: 525-533. doi: 10.1016/j.buildenv.2008.04.015
|
| [49] | Faber J, Brodzik K, Łomankiewicz D, et al. (2012) Temperature influence on air quality inside cabin of conditioned car. Combust Engines 2: 47-56. |
| [50] | Van der Wal JF, Hoogeveen AW, Wouda P (2012) The influence of temperature on the emission of volatile organic compounds from PVC flooring, carpet, and paint. Indoor Air 7: 152-159. |
| [51] | Wypych G (2013) Handbook of odors in plastic materials. ChemTec Publishing, Toronto. |
| [52] | Mitchell G, Higgitt C, Gibsion LT (2014) Emissions from polymeric materials: Characterised by thermal desorption-gas chromatography. Polym Degrad Stabil 107 Z: 328-340. |
| [53] | Yoshida T, Matsunaga I, Tomioka K, et al. (2006) Interior air pollution in automotive cabins by volatile organic compounds diffusing from interior materials: II. Influence of manufacturer, specifications and usage status on air pollution, and estimation of air pollution levels in initial phases of delivery as a new car. Indoor Built Environ 15: 445-462. |
| [54] |
Loock F, Lampe T, Bahadir M (1993) Examination of the emission of (semi-)volatile organic compounds from polymer materials used as interior trim materials in automobiles. Fresen J Anal Chem 347: 280-285. doi: 10.1007/BF00323973
|
| [55] | Khare S, Morgenstern U, Beinert C (2016) Emissions of PP-Talc-compounds during injection molding process. Proceedings of 18th Conference Odour and Emissions of Plastic Materials, Universität Kassel, Germany. |
| [56] |
Klepeis NE, Nelson WC, Ott WR, et al. (2001) The National Human Activity Pattern Survey (NHAPS): a resource for assessing exposure to environmental pollutants. J Expo Anal Environ Epid 11: 231-252. doi: 10.1038/sj.jea.7500165
|
| [57] |
Zhang GS, Li TT, Luo M, Lin JF, et al. (2008) Air pollution in the microenvironment of parked new cars. Build Environ 43: 315-319. doi: 10.1016/j.buildenv.2006.03.019
|
| [58] |
Kadiyala A, Kumar A (2012) Development and application of a methodology to identify and rank the important factors affecting in-vehicle particulate matter. J Hazard Mater 213-214: 140-146. doi: 10.1016/j.jhazmat.2012.01.072
|
| [59] |
Jo WK, Lee JH (2011) Naphthalene and benzene levels in microenvironments associated with potential exposure: new and old apartments with moth repellents, and cabins of passenger cars. Int J Environ An Ch 91: 1412-1424. doi: 10.1080/03067310903359468
|
| [60] |
Fromme H, Oddoy A, Piloty M, et al. (1998) Polycyclic aromatic hydrocarbons (PAH) and diesel engine emission (elemental carbon) inside a car and a subway train. Sci Total Environ 217: 165-173. doi: 10.1016/S0048-9697(98)00189-2
|
| [61] |
Chan AT, Chung MW (2003) Indoor – outdoor air quality relationship in vehicle: effect of driving environment and ventilation modes. Atmos Environ 37: 3795-3808. doi: 10.1016/S1352-2310(03)00466-7
|
| [62] |
Geiss O, Tirendi S, Barrero-Moreno J, et al. (2009) Investigation of volatile organic compounds and phthalates present in the cabin air of used private cars. Environ Int 35: 1188-1195. doi: 10.1016/j.envint.2009.07.016
|
| [63] |
Chan CC, Spengler JD, Özkaynak H, et al. (1991) Commuter exposures to VOCs in Boston, Massachusetts. J Air Waste Manage Assoc 41: 1594-1600. doi: 10.1080/10473289.1991.10466955
|
| [64] |
Rahman MM, Kim KH (2012) Exposure to hazardous volatile pollutants back diffusing from automobile exhaust systems. J Hazard Mater 241-242: 267-278. doi: 10.1016/j.jhazmat.2012.09.042
|
| [65] |
Chan CC, Özkaynak H, Spengler JD, et al. (1991) Driver exposure to volatile organic compounds, CO, ozone and NO2 under different driving conditions. Environ Sci Technol 25: 964-972. doi: 10.1021/es00017a021
|
| [66] |
Lee K, Sohn H, Putti K (2010) In-vehicle exposures to particulate matter and black carbon. J Air Waste Manage Assoc 60: 130-136. doi: 10.3155/1047-3289.60.2.130
|
| [67] |
Jo WK, Park KH (1999) Commuter exposure to volatile organic compounds under different driving conditions. Atmos Environ 33: 409-417. doi: 10.1016/S1352-2310(98)00201-5
|
| [68] |
Parra MA, Elustondo D, Bernejo R, et al. (2008) Exposure to volatile organic compounds (VOC) in public buses of Pamplona, Northern Spain. Sci Total Environ 404: 18-25. doi: 10.1016/j.scitotenv.2008.05.028
|
| [69] |
Leung PL, Harrison RM (1999) Roadside and in-vehicle concentrations of monoaromatic hydrocarbons. Atmos Environ 33: 191-204. doi: 10.1016/S1352-2310(98)00147-2
|
| [70] |
Faber J, Brodzik K, Gołda-Kopek A, et al. (2013) Benzene, toluene and xylenes levels in new and used vehicles of the same model. J Environ Sci 25: 2324-2330. doi: 10.1016/S1001-0742(12)60333-7
|
| [71] | Faber J, Brodzik K, Gołda-Kopek A, et al. (2014) Comparison of air pollution by VOCs inside the cabins of new vehicles. Environ Natural Resour Res 4: 155-165. |
| [72] |
Brodzik K, Faber J, Gołda-Kopek A, et al. (2014) In-vehicle VOCs composition of unconditioned, newly produced cars. J Environ Sci 26: 1052-1061. doi: 10.1016/S1001-0742(13)60459-3
|
| [73] | Faber J (2014) Air quality inside new vehicles. Investigation of air quality on the basis of volatile organic compounds presence. Saarbrücken, Germany. LAP LAMBERT Academic Publishing. |
| [74] | Faber J, Brodzik K, Gołda-Kopek A, et al. (2013) Air pollution in new vehicles as a result of VOC emission from interior materials. Pol J Environ Stud 22: 1701-1709. |
| [75] |
Buters JTM, Schober W, Gutermuth I, et al. (2007) Toxicity of parked motor vehicle indoor air. Environ Sci Technol 41: 2622-2629. doi: 10.1021/es0617901
|
| [76] |
Dor F, Le Moullec Y, Fresty B (1995) Exposure of city residents to carbon monoxide and monocyclic aromatic hydrocarbons during commuting trips in the Paris metropolitan area. J Air Waste Manage Assoc 45: 103-110. doi: 10.1080/10473289.1995.10467345
|
| [77] |
Jo WK, Song KB (2001) Exposure to volatile organic compounds for individuals with occupations associated with potential exposure to motor vehicle exhaust and/or gasoline vapor emissions. Sci Total Environ 269: 25-37. doi: 10.1016/S0048-9697(00)00774-9
|
| [78] |
Som D, Dutta C, Chatterjee A, et al. (2007) Studies on commuters' exposure to BTEX in passenger cars in Kolkata, India. Sci Total Environ 372: 426-432. doi: 10.1016/j.scitotenv.2006.09.025
|
| [79] |
Lawryk NJ, Weisel CP (1996) Concentrations of Volatile Organic Compounds in the passenger compartments of automobiles. Environ Sci Technol 30: 810-816. doi: 10.1021/es950225n
|
| [80] |
Jo WK, Lee JW (2002) In-vehicle exposure to aldehydes while commuting on real commuter routes in a Korean urban area. Environ Res A 88: 44-51. doi: 10.1006/enrs.2001.4313
|
| [81] |
Kim YM, Harrad S, Harrison RM (2001) Concentrations and sources of VOCs in urban domestic and public microenvironments. Environ Sci Technol 35: 997-1004. doi: 10.1021/es000192y
|
| [82] | Kingham S, Meaton J, Sheard A, et al. (1998) Assessment of exposure to traffic-related fumes during the journey to work. Transport Res D 3: 271-274. |
| [83] |
Tokumura M, Hatayama R, Tatsu K, et al. (2016) Car indoor air pollution by volatile organic compounds and aldehydes in Japan. AIMS Environ Sci 3: 362-381. doi: 10.3934/environsci.2016.3.362
|
| [84] |
Kuo HW, Wei HC, Liu CS, et al. (2000) Exposure to volatile organic compounds while commuting in Taichung, Taiwan. Atmos Environ 34: 3331-3336. doi: 10.1016/S1352-2310(99)00396-9
|
| [85] |
MnNabola A, Broderick BM, Gill LW (2009) The impacts of inter-vehicle spacing on in-vehicle air pollution concentrations in idling urban traffic conditions. Transport Res D 14:567-575. doi: 10.1016/j.trd.2009.08.003
|
| [86] |
Kim KH, Szulejko JE, Jo HJ, et al. (2016) Measurements of major VOCs released into the closed cabin environment of different automobiles under various engine and ventilation scenarios. Environ Pollut 215:340-346. doi: 10.1016/j.envpol.2016.05.033
|
| [87] |
Do DH, Van Langenhove H, Izuchukwu S, et al. (2014) Exposure to volatile organic compounds: comparison among different transportation modes. Atmos Environ 94: 53-62. doi: 10.1016/j.atmosenv.2014.05.019
|
| [88] | JAMA Japan, JAMA Announces voluntary guidelines for reducing vehicle cabin VOC concentrations levels. 2006. Available from: http://www.jama-english.jp/release/release/2005/050214.html |
| [89] | GB/T 27630-2011: Guideline for air quality assessment of passenger cars. Ministry of Environmental Protection, China. |
| [90] | MOLIT Notification No. 2013-549 (2013) Newly manufactured vehicle indoor air quality management standard. Ministry of Land, Infrastructure and Transport MOLIT, Korea. |
| [91] | ISO 12219-1:2012: Interior air of road vehicles. Part 1: Whole vehicle test chamber. Specification and method for the determination of volatile organic compounds in cabin interiors. |
| [92] | VIAQ-05-05, Proposal for new Mutual Resolution (M.R.X) on Vehicle Air Quality (initial version), GRPE. 2016. Available from: https://www2.unece.org/wiki/download/attachments/33849349/VIAQ-05-05%20-%20VIAQ%20Recommendation%28initial%20draft%29.docx?api=v2 |
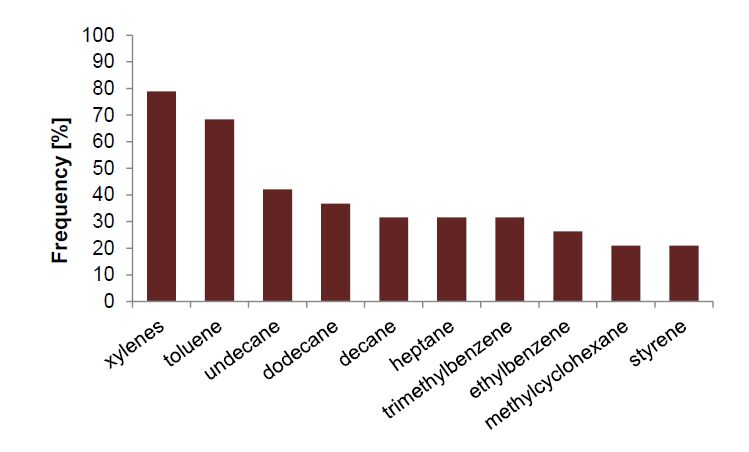
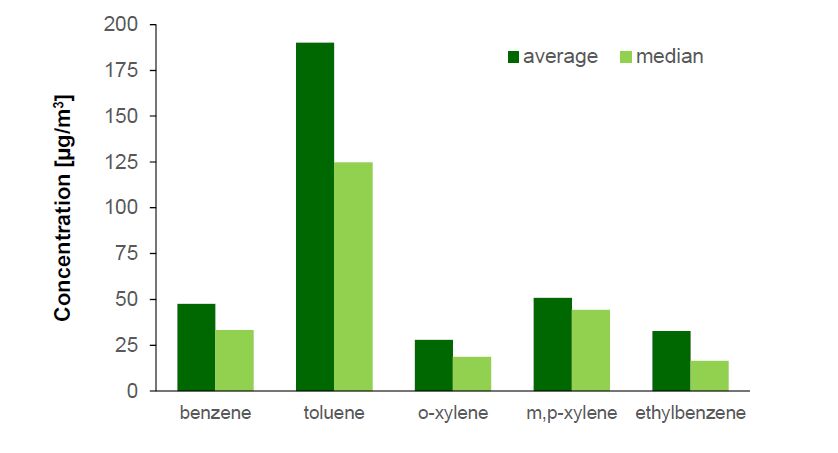
Figures(2) / Tables(5)

Joanna Faber, Krzysztof Brodzik. Air quality inside passenger cars[J]. AIMS Environmental Science, 2017, 4(1): 112-133. doi: 10.3934/environsci.2017.1.112







 DownLoad:
DownLoad: