Citation: Hassan Shirzeh, Fazel Naghdy, Philip Ciufo, Montserrat Ros. Stochastic energy balancing in substation energy management[J]. AIMS Energy, 2015, 3(4): 810-837. doi: 10.3934/energy.2015.4.810

| [1] | Clean Energy Regulator, 2014. Available from: http://www.cleanenergycouncil. org.au/policy-advocacy/reports/clean-energy-australia-report.html. |
| [2] |
Liu X (2010) Economic load dispatch constrained by wind power availability: A wait-and- see approach. Smart Grid, IEEE Transactions on 1: 347-355. doi: 10.1109/TSG.2010.2057458

|
| [3] |
Parvania M, Fotuhi-Firuzabad M (2010) Demand response scheduling by stochastic SCUC. Smart Grid, IEEE Transactions on 1: 89-98. doi: 10.1109/TSG.2010.2046430
|
| [4] |
Gong C,Wang X, Xu W, et al. (2013) Distributed real-time energy scheduling in smart grid: Stochastic model and fast optimization. Smart Grid, IEEE Transactions on 4: 1476-1489. doi: 10.1109/TSG.2013.2248399
|
| [5] |
Su S, Lu C, Chang R, et al. (2011) Distributed generation interconnection planning: A wind power case study. Smart Grid, IEEE Transactions on 2: 181-189. doi: 10.1109/TSG.2011.2105895
|
| [6] |
He M, Murugesan S, Zhang J (2013) A multi-timescale scheduling approach for stochastic reliability in smart grids with wind generation and opportunistic demand. Smart Grid, IEEE Transactions on 4: 521-529. doi: 10.1109/TSG.2012.2237421
|
| [7] | Mohd A, Ortjohann E, Schmelter A, et al. (2008) Challenges in integrating distributed energy storage systems into future smart grid. Industrial Electronics. ISIE 2008. IEEE International Symposium on: 1627-1632. |
| [8] | Dantzig G (1998) Linear Programming and Extensions, Princeton university Press. |
| [9] |
Nfaoui H, Essiarab H, Sayigh A (2004) A stochastic Markov chain model for simulating wind speed time series at Tangiers, Morocco. Renewable Energy 29: 1407-1418. doi: 10.1016/S0960-1481(03)00143-5
|
| [10] |
Hocaoglu F (2011) Stochastic approach for daily solar radiation modeling. Solar Energy 85: 278-287. doi: 10.1016/j.solener.2010.12.003
|
| [11] | Chen P, Pedersen T, Bak-Jensen B, et al. (2010) ARIMA-based time series model of stochastic wind power generation. Power Systems, IEEE Transactions on 25: 667-676. |
| [12] | Box G, Jenkins M, Reinsel G (2008) Time Series Analysis, Forecasting and Control, John Wiley & Sons Inc. |
| [13] |
Bivona S, Bonanno G, Burlon r, et al. (2011) Stochastic models for wind speed forecasting. Energy Conversion and Management 52: 1157-1165. doi: 10.1016/j.enconman.2010.09.010
|
| [14] | A Case Study of Increasing Levels of PV Penetration in an Isolated Electricity Supply Sys- tem, 2014. Available from: http://apvi.org.au/wp-content/uploads/2014/05/Carnarvon- High-PV-Penetration-Case-Study.pdf. |
| [15] |
Nguyen C, Flueck A (2012) Agent based restoration with distributed energy storage support in smart grids. Smart Grid, IEEE Transactions on 3: 1029-1038. doi: 10.1109/TSG.2012.2186833
|
| [16] |
Kodama J, Hamagami T, Shinji H, et al. (2009) Multi-agent-based autonomous power distribution network restoration using contract net protocol. Electrical Engineering in Japan 166: 56-63. doi: 10.1002/eej.20661
|
| [17] |
Lu B, Shahidehpour M (2005) Short-term scheduling of battery in a grid-connected PV/battery system. Power Systems, IEEE Transactions on 20: 1053-1061. doi: 10.1109/TPWRS.2005.846060
|
| [18] | Sortomme E, El-Sharkawi M, (2009) Optimal Power Flow for a System of Microgrids with Controllable Loads and Battery Storage. Power Systems Conference and Exposition, 2009 : 1-5. |
| [19] |
Zong Y, Kullmann D, Thavlov A, et al. (2012) Application of model predictive control for active load management in a distributed power system with high wind penetration. Smart Grid, IEEE Transactions on 3: 1055-1062. doi: 10.1109/TSG.2011.2177282
|
| [20] | Zhao Z, Wu L (2013) Impacts of high penetration wind generation and demand response on LMPs in day-ahead market. Smart Grid, IEEE Transactions on 5: 220-229. |
| [21] |
Li Y, Huang G, Xu Y, et al. (2010) Regional-scale electric power system planning under uncertainty a multistage interval-stochastic integer linear programming approach. Energy Policy 38: 475-490. doi: 10.1016/j.enpol.2009.09.038
|
| [22] |
Molderink A, Bakker V, Bosman M, et al. (2010) Management and control of domestic smart grid technology. Smart Grid, IEEE Transactions on 1: 109-119. doi: 10.1109/TSG.2010.2055904
|
| [23] |
Yu W, Liu D, Huang Y (2013) Operation Optimization Based on the Power Supply and Storage Capacity of an Active Distribution Network. Energies 6: 6423-6438. doi: 10.3390/en6126423
|
| [24] | McArthur S, Davidson E, Catterson V, et al. Multi-agent systems for power engineering applications, part ii: Technologies, standards, and tools for building multi-agent systems. Power Systems, IEEE Transactions on 22: 1753-1759. |
| [25] |
Moslehi K, Kumar R (2010) A reliability perspective of the smart grid. Smart Grid, IEEE Transactions on 1: 57-64. doi: 10.1109/TSG.2010.2046346
|
| [26] |
Garciaa-Ascanio C, Mate C (2010) Electric power demand forecasting using interval time series: A comparison between VAR and iMLP. Energy Policy 38: 715-725. doi: 10.1016/j.enpol.2009.10.007
|
| [27] |
Shirzeh H, Naghdy F, Ciufo P, et al. (2015) Balancing energy in the smart grid using distributed value function (DVF). Smart Grid, IEEE Transactions on 6: 808-818. doi: 10.1109/TSG.2014.2363844
|
| [28] | Pawlowski A, Guzman J, Rodriguez F, et al. (2010) Application of time-series methods to disturbance estimation in predictive control problems. in Industrial Electronics (ISIE), 2010 IEEE International Symposium on: 409-414. |
| [29] |
Taylor J, Menezes L, McSharry P (2006) A comparison of univariate methods for forecasting electricity demand up to a day ahead. International Journal of Forecasting 22: 1-16. doi: 10.1016/j.ijforecast.2005.06.006
|
| [30] | Munoz A, Sanchez-Ubeda E, Cruz A, et al. (2010) Short-term forecasting in power systems: A guided tour, Handbook of Power Systems II, Energy Systems, Springer Berlin Heidelberg, 129-160. |
| [31] | Pring M (2002) Technical Analysis Explained, McGraw-Hill, 2002. |
| [32] | Murphy J (1999) Technical Analysis of the Financial Markets, New York Institute of Finance. |
| [33] | Mooney C (1997) Monte Carlo Simulation, SAGE PUBLICATION. |
| [34] |
Nguyen M, Nguyen D, Yoon Y (2012) A New Battery Energy Storage Charging/Discharging Scheme for Wind Power Producers in Real-Time Markets. Energies, 5: 5439-5452. doi: 10.3390/en5125439
|
| [35] | Endeavor Energy, 2013. Available from: www.endeavourenergy.com.au/. |
| [36] | Bellifemine F, Caire G, Greenwood D (2007) Developing Multi Agent Systems with JADE, England: Wiley. |
| [37] | Department of Agriculture and Food, WA, Australia, 2014. Available from: https://www.agric.wa.gov.au/. |
| [38] | University of York, 2009. Available from: http://www.agentcontrol.co.uk/. |
| [39] | Robinson C, MendhamP, Clarke T (2010) MACSIMJX: A Tool for Enabling Agent Mod- elling with Simulink Using JADE. JoPha Journal of Pysical Agents 4: 1-7. |
| [40] | Shull F, Singer J, Sjoberg D (2008) Guide to advanced empirical software engineering, Springer. |

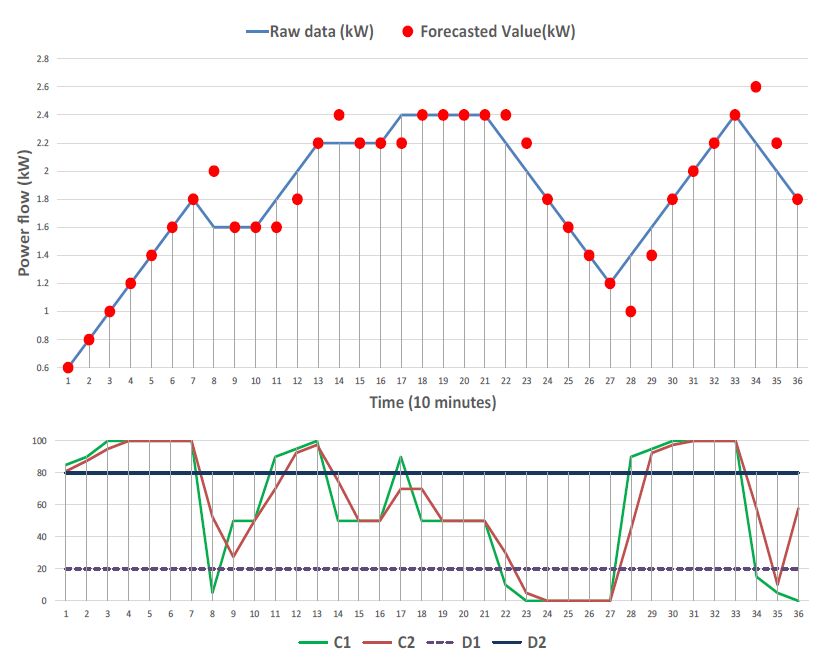
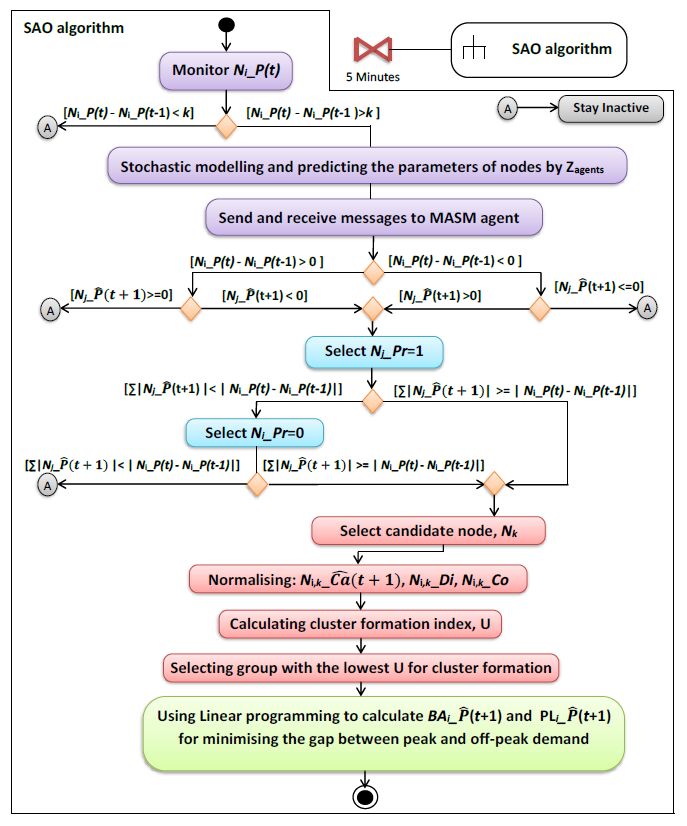
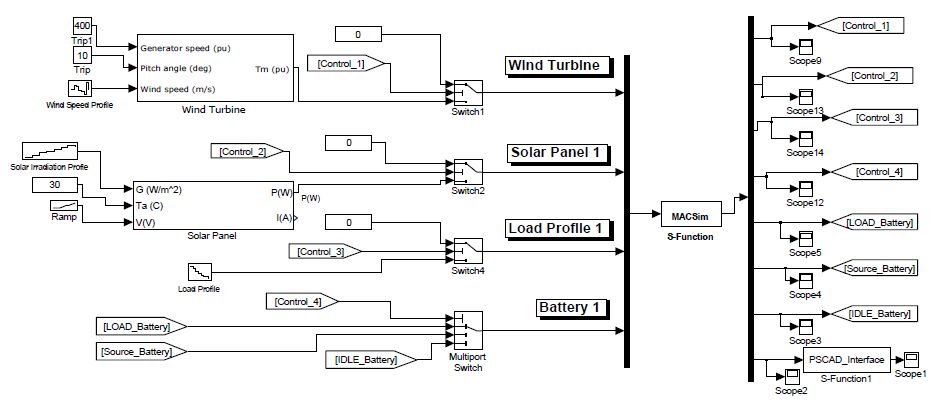
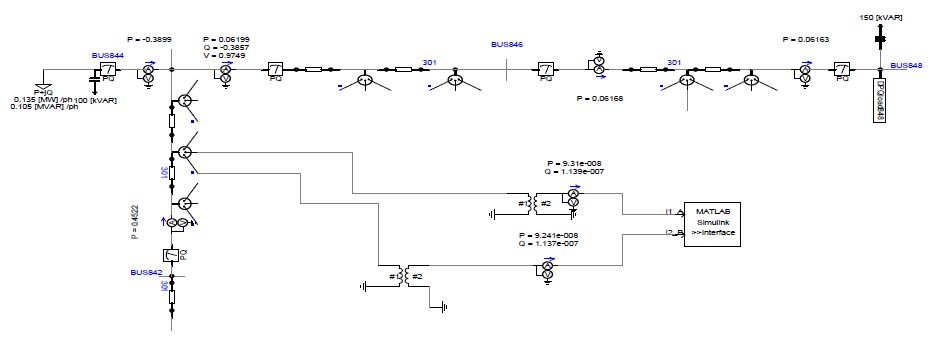



Figures(16) / Tables(3)

Hassan Shirzeh, Fazel Naghdy, Philip Ciufo, Montserrat Ros. Stochastic energy balancing in substation energy management[J]. AIMS Energy, 2015, 3(4): 810-837. doi: 10.3934/energy.2015.4.810







 DownLoad:
DownLoad: