Citation: Anastasios A. Mirisis, Anamaria Alexandrescu, Thomas J. Carew, Ashley M. Kopec. The Contribution of Spatial and Temporal Molecular Networks in the Induction of Long-term Memory and Its Underlying Synaptic Plasticity[J]. AIMS Neuroscience, 2016, 3(3): 356-384. doi: 10.3934/Neuroscience.2016.3.356

| [1] | Castellucci VF, Blumenfeld H, Goelet P, et al. (1989) Inhibitor of protein synthesis blocks long-term behavioral sensitization in the isolated gill-withdrawal reflex of Aplysia. J Neurobiol 20: 1-9. |
| [2] | Xia SZ, Feng CH, Guo AK (1998) Multiple-phase model of memory consolidation confirmed by behavioral and pharmacological analyses of operant conditioning in Drosophila. Pharmacol Biochem Behav 60: 809-816. |
| [3] | Sutton MA, Masters SE, Bagnall MW, et al. (2001) Molecular mechanisms underlying a unique intermediate phase of memory in aplysia. Neuron 31: 143-154. |
| [4] | Stough S, Shobe JL, Carew TJ (2006) Intermediate-term processes in memory formation. Curr Opin Neurobiol 16: 672-678. |
| [5] | Bailey CH, Bartsch D, Kandel ER (1996) Toward a molecular definition of long-term memory storage. Proc Natl Acad Sci U S A 93: 13445-13452. |
| [6] | Heyward FD, Sweatt JD (2015) DNA Methylation in Memory Formation: Emerging Insights. Neuroscientist 21: 475-489. |
| [7] | Alberini CM (2005) Mechanisms of memory stabilization: are consolidation and reconsolidation similar or distinct processes? Trends Neurosci 28: 51-56. |
| [8] | Pineda VV, Athos JI, Wang H, et al. (2004) Removal of G(ialpha1) constraints on adenylyl cyclase in the hippocampus enhances LTP and impairs memory formation. Neuron 41: 153-163. |
| [9] | Frey U, Morris RG (1997) Synaptic tagging and long-term potentiation. Nature 385: 533-536. |
| [10] | Sherff CM, Carew TJ (1999) Coincident induction of long-term facilitation in Aplysia: cooperativity between cell bodies and remote synapses. Science 285: 1911-1914. |
| [11] | Martin KC, Casadio A, Zhu H, et al. (1997) Synapse-specific, long-term facilitation of aplysia sensory to motor synapses: a function for local protein synthesis in memory storage. Cell 91: 927-938. |
| [12] | Giese KP, Mizuno K (2013) The roles of protein kinases in learning and memory. Learn Mem 20: 540-552. |
| [13] | Graves JD, Campbell JS, Krebs EG (1995) Protein serine/threonine kinases of the MAPK cascade. Ann N Y Acad Sci 766: 320-343. |
| [14] | Sweatt JD (2004) Mitogen-activated protein kinases in synaptic plasticity and memory. Curr Opin Neurobiol 14: 311-317. |
| [15] | Reissner KJ, Shobe JL, Carew TJ (2006) Molecular nodes in memory processing: Insights from Aplysia. Cell Moll Life Sci 63. |
| [16] | Rosen LB, Ginty DD, Weber MJ, et al. (1994) Membrane depolarization and calcium influx stimulate MEK and MAP kinase via activation of Ras. Neuron 12: 1207-1221. |
| [17] | Farnsworth CL, Freshney NW, Rosen LB, et al. (1995) Calcium activation of Ras mediated by neuronal exchange factor Ras-GRF. Nature 376: 524-527. |
| [18] | Sherrin T, Blank T, Todorovic C (2011) c-Jun N-terminal kinases in memory and synaptic plasticity. Rev Neurosci 22: 403-410. |
| [19] | Johnson GL, Lapadat R (2002) Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 298: 1911-1912. |
| [20] | Tournier C, Hess P, Yang DD, et al. (2000) Requirement of JNK for stress-induced activation of the cytochrome c-mediated death pathway. Science 288: 870-874. |
| [21] | Kyriakis JM, Banerjee P, Nikolakaki E, et al. (1994) The stress-activated protein kinase subfamily of c-Jun kinases. Nature 369: 156-160. |
| [22] | Tuvikene J, Pruunsild P, Orav E, et al. (2016) AP-1 Transcription Factors Mediate BDNF-Positive Feedback Loop in Cortical Neurons. J Neurosci 36: 1290-1305. |
| [23] | Williams JM, Beckmann AM, Mason-Parker SE, et al. (2000) Sequential increase in Egr-1 and AP-1 DNA binding activity in the dentate gyrus following the induction of long-term potentiation. Brain Res Mol Brain Res 77: 258-266. |
| [24] | Bevilaqua LR, Kerr DS, Medina JH, et al. (2003) Inhibition of hippocampal Jun N-terminal kinase enhances short-term memory but blocks long-term memory formation and retrieval of an inhibitory avoidance task. Eur J Neurosci 17: 897-902. |
| [25] | Guan Z, Kim JH, Lomvardas S, et al. (2003) p38 MAP kinase mediates both short-term and long-term synaptic depression in aplysia. J Neurosci 23: 7317-7325. |
| [26] | Alberini CM (2009) Transcription factors in long-term memory and synaptic plasticity. Physiol Rev 89: 121-145. |
| [27] | Alberini CM, Ghirardi M, Metz R, et al. (1994) C/EBP is an immediate-early gene required for the consolidation of long-term facilitation in Aplysia. Cell 76: 1099-1114. |
| [28] | Taubenfeld SM, Milekic MH, Monti B, et al. (2001) The consolidation of new but not reactivated memory requires hippocampal C/EBPbeta. Nat Neurosci 4: 813-818. |
| [29] | Murray HJ, O'Connor JJ (2003) A role for COX-2 and p38 mitogen activated protein kinase in long-term depression in the rat dentate gyrus in vitro. Neuropharmacology 44: 374-380. |
| [30] | Moult PR, Correa SA, Collingridge GL, et al. (2008) Co-activation of p38 mitogen-activated protein kinase and protein tyrosine phosphatase underlies metabotropic glutamate receptor-dependent long-term depression. J Physiol 586: 2499-2510. |
| [31] | Sanderson TM, Hogg EL, Collingridge GL, et al. (2016) Hippocampal mGluR-LTD in health and disease: focus on the p38 MAPK and ERK1/2 pathways. J Neurochem. |
| [32] | Zarubin T, Han J (2005) Activation and signaling of the p38 MAP kinase pathway. Cell Res 15: 11-18. |
| [33] | Liu RY, Zhang Y, Coughlin BL, et al. (2014) Doxorubicin attenuates serotonin-induced long-term synaptic facilitation by phosphorylation of p38 mitogen-activated protein kinase. J Neurosci 34: 13289-13300. |
| [34] | Atkins CM, Selcher JC, Petraitis JJ, et al. (1998) The MAPK cascade is required for mammalian associative learning. Nat Neurosci 1: 602-609. |
| [35] | Sharma SK, Sherff CM, Shobe J, et al. (2003) Differential role of mitogen-activated protein kinase in three distinct phases of memory for sensitization in Aplysia. J Neurosci 23: 3899-3907. |
| [36] | Ajay SM, Bhalla US (2004) A role for ERKII in synaptic selectivity on the time-scale of minutes. Eur J Neurosci 20: 2671-2680. |
| [37] | Philips GT, Ye X, Kopec AM, et al. (2013) MAPK establishes a molecular context that defines effective training patterns for long-term memory formation. J Neurosci 33: 7565-7573. |
| [38] | Ye X, Shobe JL, Sharma SK, et al. (2008) Small G proteins exhibit pattern sensitivity in MAPK activation during the induction of memory and synaptic facilitation in Aplysia. Proc Natl Acad Sci U S A 105: 20511-20516. |
| [39] | Pagani MR, Oishi K, Gelb BD, et al. (2009) The phosphatase SHP2 regulates the spacing effect for long-term memory induction. Cell 139: 186-198. |
| [40] | Shobe JL, Zhao Y, Stough S, et al. (2009) Temporal phases of activity-dependent plasticity and memory are mediated by compartmentalized routing of MAPK signaling in aplysia sensory neurons. Neuron 61: 113-125. |
| [41] | Michel M, Green CL, Eskin A, et al. (2011) PKG-mediated MAPK signaling is necessary for long-term operant memory in Aplysia. Learn Mem 18: 108-117. |
| [42] | Ye X, Marina A, Carew TJ (2012) Local synaptic integration of mitogen-activated protein kinase and protein kinase A signaling mediates intermediate-term synaptic facilitation in Aplysia. Proc Natl Acad Sci U S A 109: 18162-18167. |
| [43] | Philips GT, Tzvetkova EI, Carew TJ (2007) Transient mitogen-activated protein kinase activation is confined to a narrow temporal window required for the induction of two-trial long-term memory in Aplysia. J Neurosci 27: 13701-13705. |
| [44] | Zhang Y, Liu RY, Heberton GA, et al. (2012) Computational design of enhanced learning protocols. Nat Neurosci 15: 294-297. |
| [45] | Sweatt JD (2001) The neuronal MAP kinase cascade: a biochemical signal integration system subserving synaptic plasticity and memory. J Neurochem 76: 1-10. |
| [46] | Casar B, Pinto A, Crespo P (2009) ERK dimers and scaffold proteins: unexpected partners for a forgotten (cytoplasmic) task. Cell Cycle 8: 1007-1013. |
| [47] | Casar B, Arozarena I, Sanz-Moreno V, et al. (2009) Ras subcellular localization defines extracellular signal-regulated kinase 1 and 2 substrate specificity through distinct utilization of scaffold proteins. Mol Cell Biol 29: 1338-1353. |
| [48] | Shalin SC, Hernandez CM, Dougherty MK, et al. (2006) Kinase suppressor of Ras1 compartmentalizes hippocampal signal transduction and subserves synaptic plasticity and memory formation. Neuron 50: 765-779. |
| [49] | Morice C, Nothias F, Konig S, et al. (1999) Raf-1 and B-Raf proteins have similar regional distributions but differential subcellular localization in adult rat brain. Eur J Neurosci 11: 1995-2006. |
| [50] | Micheau J, Riedel G (1999) Protein kinases: which one is the memory molecule? Cell Mol Life Sci 55: 534-548. |
| [51] | Nguyen PV, Woo NH (2003) Regulation of hippocampal synaptic plasticity by cyclic AMP-dependent protein kinases. Prog Neurobiol 71: 401-437. |
| [52] | Baudry M, Zhu G, Liu Y, et al. (2015) Multiple cellular cascades participate in long-term potentiation and in hippocampus-dependent learning. Brain Res 1621: 73-81. |
| [53] | Kandel ER (2001) The molecular biology of memory storage: a dialog between genes and synapses. Biosci Rep 21: 565-611. |
| [54] | Kandel ER (2012) The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol Brain 5: 14. |
| [55] | Abel T, Nguyen PV, Barad M, et al. (1997) Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell 88: 615-626. |
| [56] | Hegde AN, Inokuchi K, Pei W, et al. (1997) Ubiquitin C-terminal hydrolase is an immediate-early gene essential for long-term facilitation in Aplysia. Cell 89: 115-126. |
| [57] | Muller U, Carew TJ (1998) Serotonin induces temporally and mechanistically distinct phases of persistent PKA activity in Aplysia sensory neurons. Neuron 21: 1423-1434. |
| [58] | Bacskai BJ, Hochner B, Mahaut-Smith M, et al. (1993) Spatially resolved dynamics of cAMP and protein kinase A subunits in Aplysia sensory neurons. Science 260: 222-226. |
| [59] | Kaang BK, Kandel ER, Grant SG (1993) Activation of cAMP-responsive genes by stimuli that produce long-term facilitation in Aplysia sensory neurons. Neuron 10: 427-435. |
| [60] | Dash PK, Hochner B, Kandel ER (1990) Injection of the cAMP-responsive element into the nucleus of Aplysia sensory neurons blocks long-term facilitation. Nature 345: 718-721. |
| [61] | Pu L, Bao GB, Xu NJ, et al. (2002) Hippocampal long-term potentiation is reduced by chronic opiate treatment and can be restored by re-exposure to opiates. J Neurosci 22: 1914-1921. |
| [62] | Bourtchuladze R, Frenguelli B, Blendy J, et al. (1994) Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell 79: 59-68. |
| [63] | Bauman AL, Goehring AS, Scott JD (2004) Orchestration of synaptic plasticity through AKAP signaling complexes. Neuropharmacology 46: 299-310. |
| [64] | Sanderson JL, Dell'Acqua ML (2011) AKAP signaling complexes in regulation of excitatory synaptic plasticity. Neuroscientist 17: 321-336. |
| [65] | Huang T, McDonough CB, Abel T (2006) Compartmentalized PKA signaling events are required for synaptic tagging and capture during hippocampal late-phase long-term potentiation. Eur J Cell Biol 85: 635-642. |
| [66] | Guan Z, Giustetto M, Lomvardas S, et al. (2002) Integration of long-term-memory-related synaptic plasticity involves bidirectional regulation of gene expression and chromatin structure. Cell 111: 483-493. |
| [67] | Bartsch D, Ghirardi M, Skehel PA, et al. (1995) Aplysia CREB2 represses long-term facilitation: relief of repression converts transient facilitation into long-term functional and structural change. Cell 83: 979-992. |
| [68] | Abel T, Martin KC, Bartsch D, et al. (1998) Memory suppressor genes: inhibitory constraints on the storage of long-term memory. Science 279: 338-341. |
| [69] | Hai T, Hartman MG (2001) The molecular biology and nomenclature of the activating transcription factor/cAMP responsive element binding family of transcription factors: activating transcription factor proteins and homeostasis. Gene 273: 1-11. |
| [70] | Schoch S, Cibelli G, Magin A, et al. (2001) Modular structure of cAMP response element binding protein 2 (CREB2). Neurochem Int 38: 601-608. |
| [71] | Pasini S, Corona C, Liu J, et al. (2015) Specific downregulation of hippocampal ATF4 reveals a necessary role in synaptic plasticity and memory. Cell Rep 11: 183-191. |
| [72] | Soda T, Frank C, Ishizuka K, et al. (2013) DISC1-ATF4 transcriptional repression complex: dual regulation of the cAMP-PDE4 cascade by DISC1. Mol Psychiatry 18: 898-908. |
| [73] | Roberson ED, English JD, Adams JP, et al. (1999) The mitogen-activated protein kinase cascade couples PKA and PKC to cAMP response element binding protein phosphorylation in area CA1 of hippocampus. J Neurosci 19: 4337-4348. |
| [74] | Martin KC, Michael D, Rose JC, et al. (1997) MAP kinase translocates into the nucleus of the presynaptic cell and is required for long-term facilitation in Aplysia. Neuron 18: 899-912. |
| [75] | Impey S, Obrietan K, Wong ST, et al. (1998) Cross talk between ERK and PKA is required for Ca2+ stimulation of CREB-dependent transcription and ERK nuclear translocation. Neuron 21: 869-883. |
| [76] | Rosenkranz JA, Frick A, Johnston D (2009) Kinase-dependent modification of dendritic excitability after long-term potentiation. J Physiol 587: 115-125. |
| [77] | Quevedo J, Vianna MR, Roesler R, et al. (2005) Pretraining but not preexposure to the task apparatus prevents the memory impairment induced by blockade of protein synthesis, PKA or MAP kinase in rats. Neurochem Res 30: 61-67. |
| [78] | Wu GY, Deisseroth K, Tsien RW (2001) Activity-dependent CREB phosphorylation: convergence of a fast, sensitive calmodulin kinase pathway and a slow, less sensitive mitogen-activated protein kinase pathway. Proc Natl Acad Sci U S A 98: 2808-2813. |
| [79] | Wayman GA, Lee YS, Tokumitsu H, et al. (2008) Calmodulin-kinases: modulators of neuronal development and plasticity. Neuron 59: 914-931. |
| [80] | Kelly MT, Crary JF, Sacktor TC (2007) Regulation of protein kinase Mzeta synthesis by multiple kinases in long-term potentiation. J Neurosci 27: 3439-3444. |
| [81] | Newton AC (1995) Protein kinase C: structure, function, and regulation. J Biol Chem 270: 28495-28498. |
| [82] | Tsokas P, Ma T, Iyengar R, et al. (2007) Mitogen-activated protein kinase upregulates the dendritic translation machinery in long-term potentiation by controlling the mammalian target of rapamycin pathway. J Neurosci 27: 5885-5894. |
| [83] | Cammalleri M, Lutjens R, Berton F, et al. (2003) Time-restricted role for dendritic activation of the mTOR-p70S6K pathway in the induction of late-phase long-term potentiation in the CA1. Proc Natl Acad Sci U S A 100: 14368-14373. |
| [84] | Kelleher RJ, 3rd, Govindarajan A, Tonegawa S (2004) Translational regulatory mechanisms in persistent forms of synaptic plasticity. Neuron 44: 59-73. |
| [85] | Hoeffer CA, Klann E (2010) mTOR signaling: at the crossroads of plasticity, memory and disease. Trends Neurosci 33: 67-75. |
| [86] | Hayer A, Bhalla US (2005) Molecular switches at the synapse emerge from receptor and kinase traffic. PLoS Comput Biol 1: 137-154. |
| [87] | Smolen P, Baxter DA, Byrne JH (2006) A model of the roles of essential kinases in the induction and expression of late long-term potentiation. Biophys J 90: 2760-2775. |
| [88] | Jalil SJ, Sacktor TC, Shouval HZ (2015) Atypical PKCs in memory maintenance: the roles of feedback and redundancy. Learn Mem 22: 344-353. |
| [89] | Heerssen HM, Segal RA (2002) Location, location, location: a spatial view of neurotrophin signal transduction. Trends Neurosci 25: 160-165. |
| [90] | Dailey L, Ambrosetti D, Mansukhani A, et al. (2005) Mechanisms underlying differential responses to FGF signaling. Cytokine Growth Factor Rev 16: 233-247. |
| [91] | Ramel MC, Hill CS (2012) Spatial regulation of BMP activity. FEBS Lett 586: 1929-1941. |
| [92] | Kopec AM, Carew TJ (2013) Growth factor signaling and memory formation: temporal and spatial integration of a molecular network. Learn Mem 20: 531-539. |
| [93] | Cohen S, Levi-Montalcini R, Hamburger V (1954) A nerve growth-stimulating factor isolated from sarcomas 37 and 180. Proc Natl Acad Sci U S A 40: 1014-1018. |
| [94] | Levi-Montalcini R, Skaper SD, Dal Toso R, et al. (1996) Nerve growth factor: from neurotrophin to neurokine. Trends Neurosci 19: 514-520. |
| [95] | Edelmann E, Lessmann V, Brigadski T (2014) Pre- and postsynaptic twists in BDNF secretion and action in synaptic plasticity. Neuropharmacology 76 Pt C: 610-627. |
| [96] | Lessmann V, Gottmann K, Malcangio M (2003) Neurotrophin secretion: current facts and future prospects. Prog Neurobiol 69: 341-374. |
| [97] | Tongiorgi E (2008) Activity-dependent expression of brain-derived neurotrophic factor in dendrites: facts and open questions. Neurosci Res 61: 335-346. |
| [98] | Conner JM, Lauterborn JC, Yan Q, et al. (1997) Distribution of brain-derived neurotrophic factor (BDNF) protein and mRNA in the normal adult rat CNS: evidence for anterograde axonal transport. J Neurosci 17: 2295-2313. |
| [99] | Smith MA, Zhang LX, Lyons WE, et al. (1997) Anterograde transport of endogenous brain-derived neurotrophic factor in hippocampal mossy fibers. Neuroreport 8: 1829-1834. |
| [100] | Yan Q, Rosenfeld RD, Matheson CR, et al. (1997) Expression of brain-derived neurotrophic factor protein in the adult rat central nervous system. Neuroscience 78: 431-448. |
| [101] | Salio C, Averill S, Priestley JV, et al. (2007) Costorage of BDNF and neuropeptides within individual dense-core vesicles in central and peripheral neurons. Dev Neurobiol 67: 326-338. |
| [102] | Dieni S, Matsumoto T, Dekkers M, et al. (2012) BDNF and its pro-peptide are stored in presynaptic dense core vesicles in brain neurons. J Cell Biol 196: 775-788. |
| [103] | Altar CA, Cai N, Bliven T, et al. (1997) Anterograde transport of brain-derived neurotrophic factor and its role in the brain. Nature 389: 856-860. |
| [104] | Wetmore C, Cao YH, Pettersson RF, et al. (1991) Brain-derived neurotrophic factor: subcellular compartmentalization and interneuronal transfer as visualized with anti-peptide antibodies. Proc Natl Acad Sci U S A 88: 9843-9847. |
| [105] | Aoki C, Wu K, Elste A, et al. (2000) Localization of brain-derived neurotrophic factor and TrkB receptors to postsynaptic densities of adult rat cerebral cortex. J Neurosci Res 59: 454-463. |
| [106] | Tongiorgi E, Armellin M, Giulianini PG, et al. (2004) Brain-derived neurotrophic factor mRNA and protein are targeted to discrete dendritic laminas by events that trigger epileptogenesis. J Neurosci 24: 6842-6852. |
| [107] | An JJ, Gharami K, Liao GY, et al. (2008) Distinct role of long 3' UTR BDNF mRNA in spine morphology and synaptic plasticity in hippocampal neurons. Cell 134: 175-187. |
| [108] | DiStefano PS, Friedman B, Radziejewski C, et al. (1992) The neurotrophins BDNF, NT-3, and NGF display distinct patterns of retrograde axonal transport in peripheral and central neurons. Neuron 8: 983-993. |
| [109] | Zweifel LS, Kuruvilla R, Ginty DD (2005) Functions and mechanisms of retrograde neurotrophin signalling. Nat Rev Neurosci 6: 615-625. |
| [110] | Baulida J, Kraus MH, Alimandi M, et al. (1996) All ErbB receptors other than the epidermal growth factor receptor are endocytosis impaired. Journal of Biological Chemistry 271: 5251-5257. |
| [111] | Lau MM, Stewart CE, Liu Z, et al. (1994) Loss of the imprinted IGF2/cation-independent mannose 6-phosphate receptor results in fetal overgrowth and perinatal lethality. Genes Dev 8: 2953-2963. |
| [112] | Hawkes C, Kar S (2004) The insulin-like growth factor-II/mannose-6-phosphate receptor: structure, distribution and function in the central nervous system. Brain Res Brain Res Rev 44: 117-140. |
| [113] | Bao J, Alroy I, Waterman H, et al. (2000) Threonine phosphorylation diverts internalized epidermal growth factor receptors from a degradative pathway to the recycling endosome. Journal of Biological Chemistry 275: 26178-26186. |
| [114] | Lu B (2003) BDNF and activity-dependent synaptic modulation. Learn Mem 10: 86-98. |
| [115] | Lessmann V, Brigadski T (2009) Mechanisms, locations, and kinetics of synaptic BDNF secretion: an update. Neurosci Res 65: 11-22. |
| [116] | Zafra F, Hengerer B, Leibrock J, et al. (1990) Activity dependent regulation of BDNF and NGF mRNAs in the rat hippocampus is mediated by non-NMDA glutamate receptors. Embo j 9: 3545-3550. |
| [117] | Blochl A, Thoenen H (1995) Characterization of nerve growth factor (NGF) release from hippocampal neurons: evidence for a constitutive and an unconventional sodium-dependent regulated pathway. Eur J Neurosci 7: 1220-1228. |
| [118] | Blochl A, Thoenen H (1996) Localization of cellular storage compartments and sites of constitutive and activity-dependent release of nerve growth factor (NGF) in primary cultures of hippocampal neurons. Mol Cell Neurosci 7: 173-190. |
| [119] | Goodman LJ, Valverde J, Lim F, et al. (1996) Regulated release and polarized localization of brain-derived neurotrophic factor in hippocampal neurons. Mol Cell Neurosci 7: 222-238. |
| [120] | Boukhaddaoui H, Sieso V, Scamps F, et al. (2001) An activity-dependent neurotrophin-3 autocrine loop regulates the phenotype of developing hippocampal pyramidal neurons before target contact. J Neurosci 21: 8789-8797. |
| [121] | Patterson SL, Grover LM, Schwartzkroin PA, et al. (1992) Neurotrophin expression in rat hippocampal slices: a stimulus paradigm inducing LTP in CA1 evokes increases in BDNF and NT-3 mRNAs. Neuron 9: 1081-1088. |
| [122] | Canossa M, Griesbeck O, Berninger B, et al. (1997) Neurotrophin release by neurotrophins: implications for activity-dependent neuronal plasticity. Proc Natl Acad Sci U S A 94: 13279-13286. |
| [123] | Kruttgen A, Moller JC, Heymach JV, Jr., et al. (1998) Neurotrophins induce release of neurotrophins by the regulated secretory pathway. Proc Natl Acad Sci U S A 95: 9614-9619. |
| [124] | Bambah-Mukku D, Travaglia A, Chen DY, et al. (2014) A Positive Autoregulatory BDNF Feedback Loop via C/EBPbeta Mediates Hippocampal Memory Consolidation. J Neurosci 34: 12547-12559. |
| [125] | Kim HG, Wang T, Olafsson P, et al. (1994) Neurotrophin 3 potentiates neuronal activity and inhibits gamma-aminobutyratergic synaptic transmission in cortical neurons. Proc Natl Acad Sci U S A 91: 12341-12345. |
| [126] | Carmignoto G, Pizzorusso T, Tia S, et al. (1997) Brain-derived neurotrophic factor and nerve growth factor potentiate excitatory synaptic transmission in the rat visual cortex. J Physiol 498 ( Pt 1): 153-164. |
| [127] | Lessmann V, Gottmann K, Heumann R (1994) BDNF and NT-4/5 enhance glutamatergic synaptic transmission in cultured hippocampal neurones. Neuroreport 6: 21-25. |
| [128] | Kang H, Schuman EM (1995) Long-lasting neurotrophin-induced enhancement of synaptic transmission in the adult hippocampus. Science 267: 1658-1662. |
| [129] | Levine ES, Crozier RA, Black IB, et al. (1998) Brain-derived neurotrophic factor modulates hippocampal synaptic transmission by increasing N-methyl-D-aspartic acid receptor activity. Proc Natl Acad Sci U S A 95: 10235-10239. |
| [130] | Levine ES, Dreyfus CF, Black IB, et al. (1995) Brain-derived neurotrophic factor rapidly enhances synaptic transmission in hippocampal neurons via postsynaptic tyrosine kinase receptors. Proc Natl Acad Sci U S A 92: 8074-8077. |
| [131] | Tanaka T, Saito H, Matsuki N (1997) Inhibition of GABAA synaptic responses by brain-derived neurotrophic factor (BDNF) in rat hippocampus. J Neurosci 17: 2959-2966. |
| [132] | Chen DY, Bambah-Mukku D, Pollonini G, et al. (2012) Glucocorticoid receptors recruit the CaMKIIalpha-BDNF-CREB pathways to mediate memory consolidation. Nat Neurosci 15: 1707-1714. |
| [133] | Kanhema T, Dagestad G, Panja D, et al. (2006) Dual regulation of translation initiation and peptide chain elongation during BDNF-induced LTP in vivo: evidence for compartment-specific translation control. J Neurochem 99: 1328-1337. |
| [134] | Chen DY, Stern SA, Garcia-Osta A, et al. (2011) A critical role for IGF-II in memory consolidation and enhancement. Nature 469: 491-497. |
| [135] | Woolf NJ, Milov AM, Schweitzer ES, et al. (2001) Elevation of nerve growth factor and antisense knockdown of TrkA receptor during contextual memory consolidation. J Neurosci 21: 1047-1055. |
| [136] | Ageta H, Ikegami S, Miura M, et al. (2010) Activin plays a key role in the maintenance of long-term memory and late-LTP. Learning & Memory 17: 176-185. |
| [137] | Bekinschtein P, Cammarota M, Igaz LM, et al. (2007) Persistence of long-term memory storage requires a late protein synthesis- and BDNF- dependent phase in the hippocampus. Neuron 53: 261-277. |
| [138] | Bekinschtein P, Cammarota M, Katche C, et al. (2008) BDNF is essential to promote persistence of long-term memory storage. Proc Natl Acad Sci U S A 105: 2711-2716. |
| [139] | Matus A (2000) Actin-based plasticity in dendritic spines. Science 290: 754-758. |
| [140] | Jaworski J, Kapitein LC, Gouveia SM, et al. (2009) Dynamic microtubules regulate dendritic spine morphology and synaptic plasticity. Neuron 61: 85-100. |
| [141] | Tsvetkov A, Popov S (2007) Analysis of tubulin transport in nerve processes. Methods Mol Med 137: 161-173. |
| [142] | Hirokawa N (1998) Kinesin and dynein superfamily proteins and the mechanism of organelle transport. Science 279: 519-526. |
| [143] | Goldstein LS, Philp AV (1999) The road less traveled: emerging principles of kinesin motor utilization. Annu Rev Cell Dev Biol 15: 141-183. |
| [144] | Woehlke G, Schliwa M (2000) Walking on two heads: the many talents of kinesin. Nat Rev Mol Cell Biol 1: 50-58. |
| [145] | Merriam EB, Millette M, Lumbard DC, et al. (2013) Synaptic regulation of microtubule dynamics in dendritic spines by calcium, F-actin, and drebrin. J Neurosci 33: 16471-16482. |
| [146] | Puthanveettil SV, Monje FJ, Miniaci MC, et al. (2008) A new component in synaptic plasticity: upregulation of kinesin in the neurons of the gill-withdrawal reflex. Cell 135: 960-973. |
| [147] | Pinsker HM, Hening WA, Carew TJ, et al. (1973) Long-term sensitization of a defensive withdrawal reflex in Aplysia. Science 182: 1039-1042. |
| [148] | Marinesco S, Carew TJ (2002) Serotonin release evoked by tail nerve stimulation in the CNS of aplysia: characterization and relationship to heterosynaptic plasticity. J Neurosci 22: 2299-2312. |
| [149] | Philips GT, Sherff CM, Menges SA, et al. (2011) The tail-elicited tail withdrawal reflex of Aplysia is mediated centrally at tail sensory-motor synapses and exhibits sensitization across multiple temporal domains. Learn Mem 18: 272-282. |
| [150] | Hawkins RD, Kandel ER, Bailey CH (2006) Molecular mechanisms of memory storage in Aplysia. Biol Bull 210: 174-191. |
| [151] | Sutton MA, Ide J, Masters SE, et al. (2002) Interaction between amount and pattern of training in the induction of intermediate- and long-term memory for sensitization in aplysia. Learn Mem 9: 29-40. |
| [152] | Mauelshagen J, Sherff CM, Carew TJ (1998) Differential induction of long-term synaptic facilitation by spaced and massed applications of serotonin at sensory neuron synapses of Aplysia californica. Learn Mem 5: 246-256. |
| [153] | Philips GT, Kopec AM, Carew TJ (2013) Pattern and predictability in memory formation: From molecular mechanisms to clinical relevance. Neurobiol Learn Mem. |
| [154] | Shobe J, Philips GT, Carew TJ (2016) Transforming growth factor beta recruits persistent MAPK signaling to regulate long-term memory consolidation in Aplysia californica. Learn Mem 23: 182-188. |
| [155] | Sharma SK, Sherff CM, Stough S, et al. (2006) A tropomyosin-related kinase B ligand is required for ERK activation, long-term synaptic facilitation, and long-term memory in aplysia. Proc Natl Acad Sci U S A 103: 14206-14210. |
| [156] | Kassabov SR, Choi YB, Karl KA, et al. (2013) A Single Aplysia Neurotrophin Mediates Synaptic Facilitation via Differentially Processed Isoforms. Cell Rep 3: 1213-1227. |
| [157] | Chin J, Angers A, Cleary LJ, et al. (1999) TGF-beta 1 in Aplysia: Role in long-term changes in the excitability of sensory neurons and distribution of T beta R-II-like immunoreactivity. Learning & Memory 6: 317-330. |
| [158] | Chin J, Angers A, Cleary LJ, et al. (2002) Transforming growth factor beta1 alters synapsin distribution and modulates synaptic depression in Aplysia. J Neurosci 22: RC220. |
| [159] | Chin J, Liu RY, Cleary LJ, et al. (2006) TGF-beta1-induced long-term changes in neuronal excitability in aplysia sensory neurons depend on MAPK. J Neurophysiol 95: 3286-3290. |
| [160] | Zhang F, Endo S, Cleary LJ, et al. (1997) Role of transforming growth factor-beta in long-term synaptic facilitation in Aplysia. Science 275: 1318-1320. |
| [161] | Kopec AM, Philips GT, Carew TJ (2015) Distinct Growth Factor Families Are Recruited in Unique Spatiotemporal Domains during Long-Term Memory Formation in Aplysia californica. Neuron 86: 1228-1239. |
| [162] | Yim SJ, Lee YS, Lee JA, et al. (2006) Regulation of ApC/EBP mRNA by the Aplysia AU-rich element-binding protein, ApELAV, and its effects on 5-hydroxytryptamine-induced long-term facilitation. J Neurochem 98: 420-429. |
| [163] | Glisovic T, Bachorik JL, Yong J, et al. (2008) RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett 582: 1977-1986. |
| [164] | Sanchez-Diaz P, Penalva LO (2006) Post-transcription meets post-genomic: the saga of RNA binding proteins in a new era. RNA Biol 3: 101-109. |
| [165] | Wyers F, Rougemaille M, Badis G, et al. (2005) Cryptic pol II transcripts are degraded by a nuclear quality control pathway involving a new poly(A) polymerase. Cell 121: 725-737. |
| [166] | Doller A, Schlepckow K, Schwalbe H, et al. (2010) Tandem phosphorylation of serines 221 and 318 by protein kinase Cdelta coordinates mRNA binding and nucleocytoplasmic shuttling of HuR. Mol Cell Biol 30: 1397-1410. |
| [167] | Lafarga V, Cuadrado A, Lopez de Silanes I, et al. (2009) p38 Mitogen-activated protein kinase- and HuR-dependent stabilization of p21(Cip1) mRNA mediates the G(1)/S checkpoint. Mol Cell Biol 29: 4341-4351. |
| [168] | Wang W, Fan J, Yang X, et al. (2002) AMP-activated kinase regulates cytoplasmic HuR. Mol Cell Biol 22: 3425-3436. |
| [169] | Bakken TE, Miller JA, Ding SL, et al. (2016) A comprehensive transcriptional map of primate brain development. Nature 535: 367-375. |
| [170] | Yuzwa SA, Yang G, Borrett MJ, et al. (2016) Proneurogenic Ligands Defined by Modeling Developing Cortex Growth Factor Communication Networks. Neuron. |
| [171] | Pollak DD, Minh BQ, Cicvaric A, et al. (2014) A novel fibroblast growth factor receptor family member promotes neuronal outgrowth and synaptic plasticity in aplysia. Amino Acids 46: 2477-2488. |
| [172] | Pu L, Kopec AM, Boyle HD, et al. (2014) A novel cysteine-rich neurotrophic factor in Aplysia facilitates growth, MAPK activation, and long-term synaptic facilitation. Learn Mem 21: 215-222. |
| [173] | Brunet JF, Shapiro E, Foster SA, et al. (1991) Identification of a peptide specific for Aplysia sensory neurons by PCR-based differential screening. Science 252: 856-859. |
| [174] | Phillips HS, Hains JM, Armanini M, et al. (1991) BDNF mRNA is decreased in the hippocampus of individuals with Alzheimer's disease. Neuron 7: 695-702. |
| [175] | Peress NS, Perillo E (1995) Differential expression of TGF-beta 1, 2 and 3 isotypes in Alzheimer's disease: a comparative immunohistochemical study with cerebral infarction, aged human and mouse control brains. J Neuropathol Exp Neurol 54: 802-811. |
| [176] | Lim YY, Villemagne VL, Laws SM, et al. (2014) APOE and BDNF polymorphisms moderate amyloid beta-related cognitive decline in preclinical Alzheimer's disease. Mol Psychiatry. |
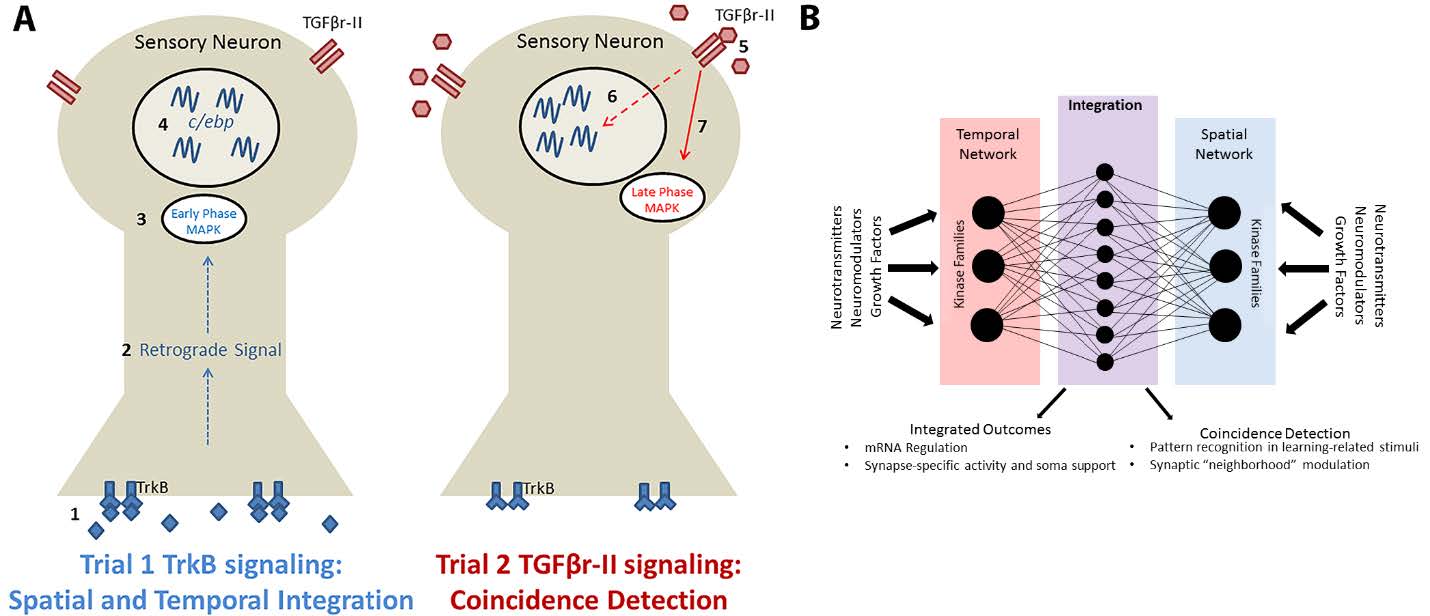
Figures(1)

Anastasios A. Mirisis, Anamaria Alexandrescu, Thomas J. Carew, Ashley M. Kopec. The Contribution of Spatial and Temporal Molecular Networks in the Induction of Long-term Memory and Its Underlying Synaptic Plasticity[J]. AIMS Neuroscience, 2016, 3(3): 356-384. doi: 10.3934/Neuroscience.2016.3.356







 DownLoad:
DownLoad: