We study numerically a coagulation-fragmentation model derived by Niwa [
Citation: Pierre Degond, Maximilian Engel. Numerical approximation of a coagulation-fragmentation model for animal group size statistics[J]. Networks and Heterogeneous Media, 2017, 12(2): 217-243. doi: 10.3934/nhm.2017009
We study numerically a coagulation-fragmentation model derived by Niwa [

| [1] |
Possible universality in the size distribution of fish schools. Phys. Rev. E (1995) 51: 5220-5223. 
|
| [2] |
Scaling in animal group-size distributions. Proc. Natl. Acad. Sci. USA (1999) 96: 4472-4477.
|
| [3] |
Convergence of a finite volume scheme for coagulation-fragmentation equations. Comm. Math. Sciences (2008) 77: 851-882.
|
| [4] | (2011) Functional Analysis, Sobolev Spaces and Partial Differential Equations. New York: Springer-Verlag. |
| [5] |
Coagulation-fragmentation model for animal group-size statistics. J. Nonlinear Sci. (2017) 27: 379-424.
|
| [6] |
Numerical simulation of the Smoluchowski coagulation equation. SIAM J. Sci. Comput. (2004) 25: 2004-2028.
|
| [7] |
A finite volume preserving scheme on nonuniform meshes and for multidimensional coalescence. SIAM J. Sci. Comput. (2012) 34: B840-B860.
|
| [8] |
The steady-state distributions of coagulation-fragmentation processes. J. Math. Biol. (1998) 37: 1-27.
|
| [9] |
The dynamics of group formations. Math. Biosc. (1995) 128: 243-264.
|
| [10] |
An accurate and efficient discrete formulation of aggregation population balance equation. Kinet. Relat. Models (2016) 9: 373-391.
|
| [11] |
Moment preserving finite volume schemes for solving population balance equations incorporating aggregation, breakage, growth and source terms. Math. Models Methods Appl. Sci. (2013) 23: 1235-1273.
|
| [12] |
A first principles derivation of animal group size distributions. J. Theoret. Biol. (2011) 283: 35-43.
|
| [13] |
Efficient solution of population balance equations with discontinuities by finite elements. Chem. Eng. Sci. (2002) 57: 1107-1119.
|
| [14] |
A finite element analysis of the steady state population balance equation for particulate systems: Aggregation and growth. Comput. Chem. Eng. (1996) 20: 261-266.
|
| [15] |
Mathematical model for the size distributions of fish schools. Comp. Math. Appl. (1996) 32: 79-88.
|
| [16] |
School size statistics of fish. J. Theoret. Biol. (1998) 195: 351-361.
|
| [17] |
Power-Law versus exponential distributions of animal group sizes. J. Theoret. Biol. (2003) 224: 451-457.
|
| [18] |
Space-irrelevant scaling law for fish school sizes. J. Theoret. Biol. (2004) 228: 347-357.
|
| [19] | Dynamical aspects of animal grouping: Swarms, schools, rocks, and herds. Adv. Biophys. (1986) 22: 1-94. |
| [20] |
Finite-element scheme for solution of the dynamic population balance equation. AIChE Journal (2003) 49: 1127-1139.
|
| [21] |
Population balances for particulate processes-a volume approach. Chem. Eng. Sci. (2002) 57: 2287-2303.
|
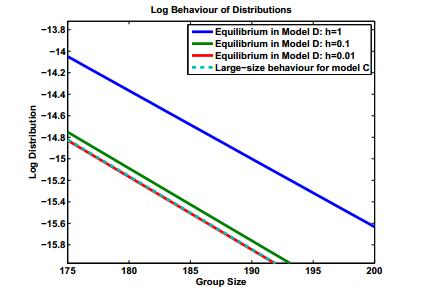
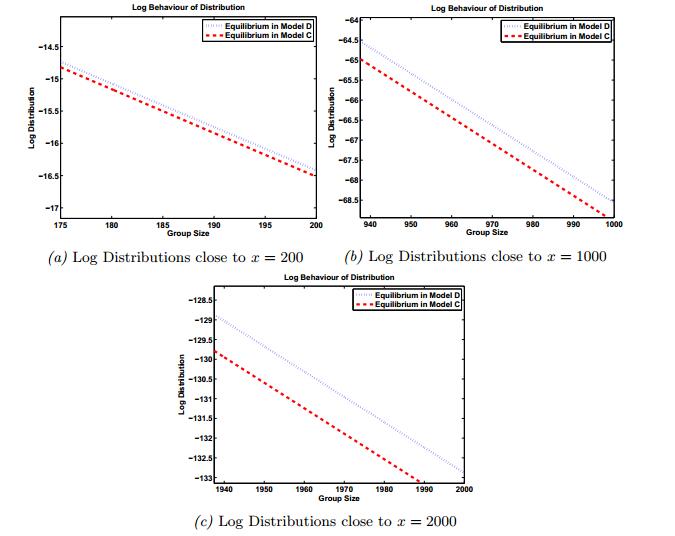
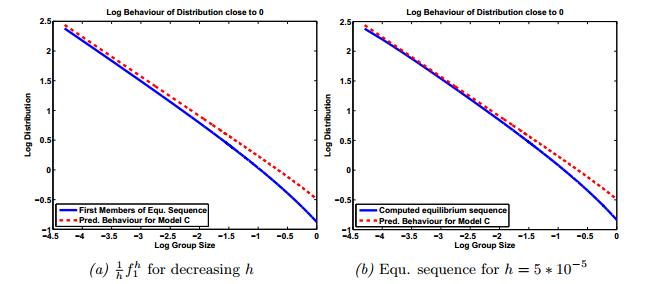
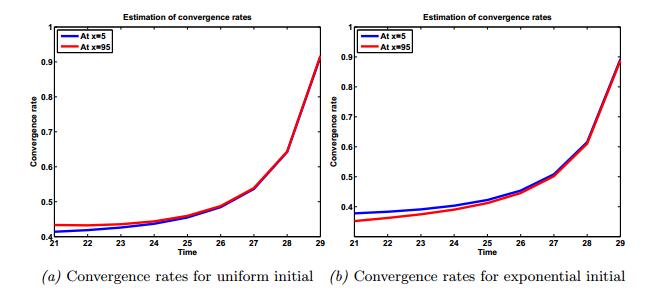
Figures(8) / Tables(2)
Pierre Degond, Maximilian Engel. Numerical approximation of a coagulation-fragmentation model for animal group size statistics[J]. Networks and Heterogeneous Media, 2017, 12(2): 217-243. doi: 10.3934/nhm.2017009







 DownLoad:
DownLoad: