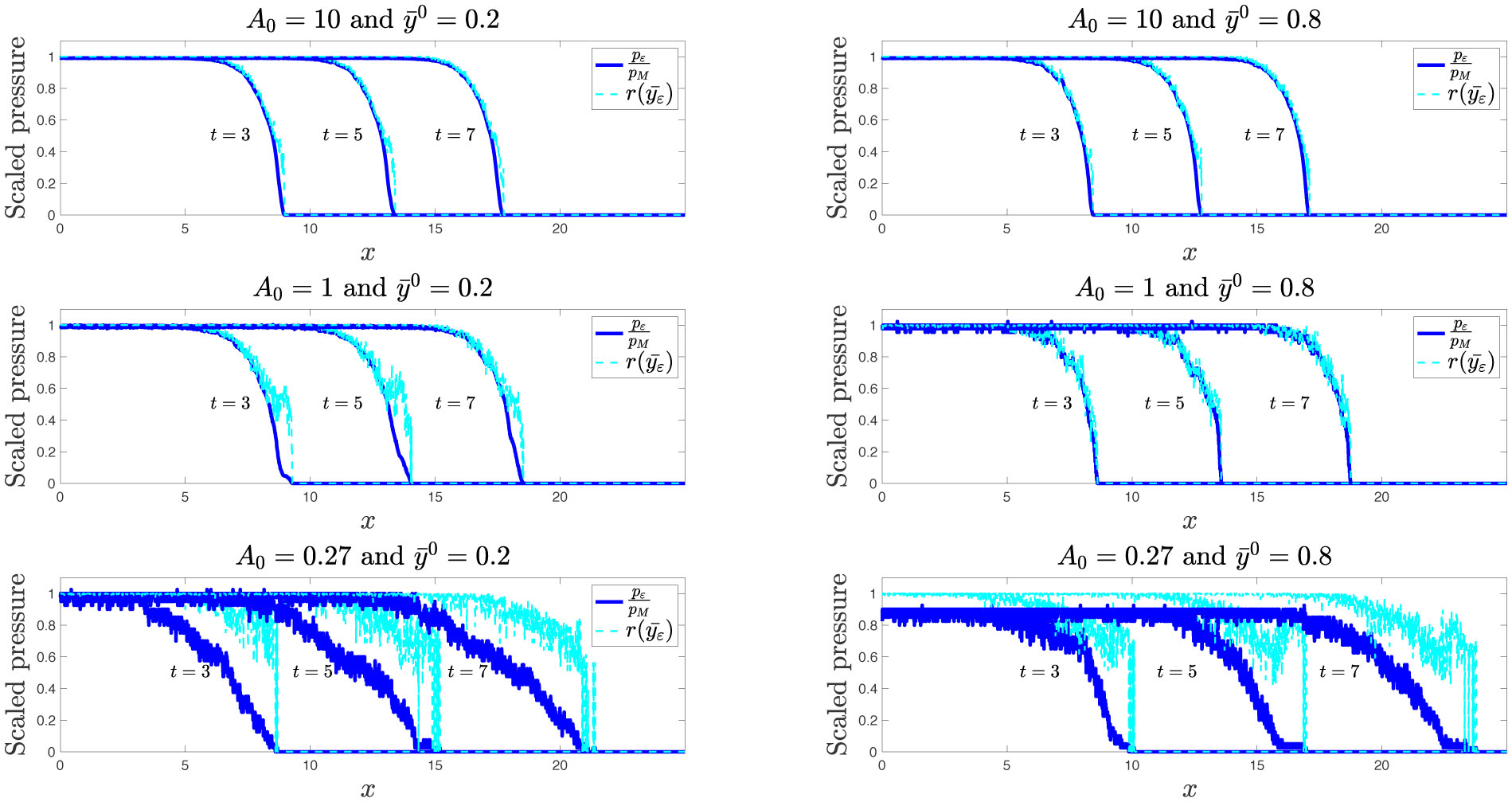
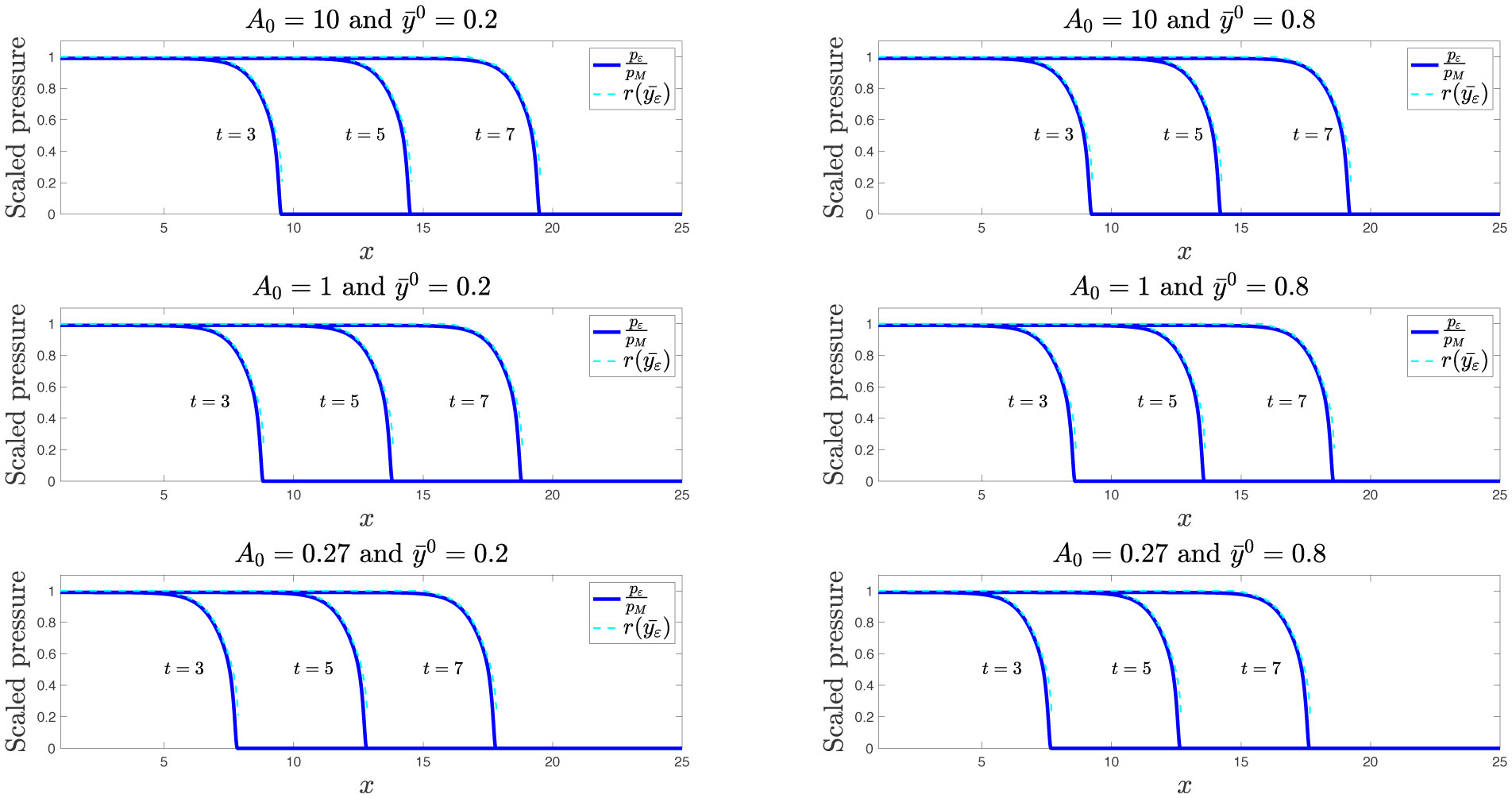
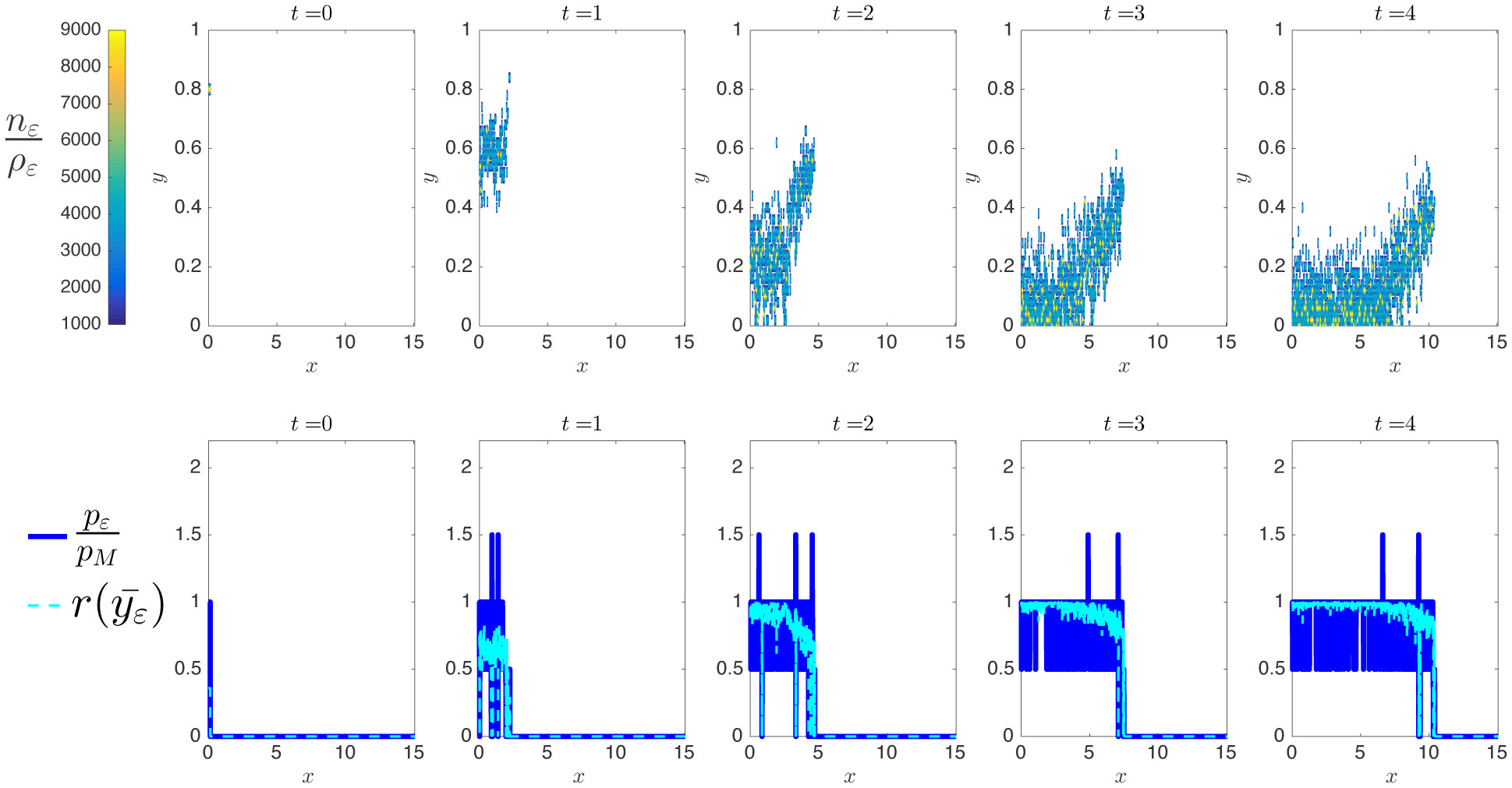
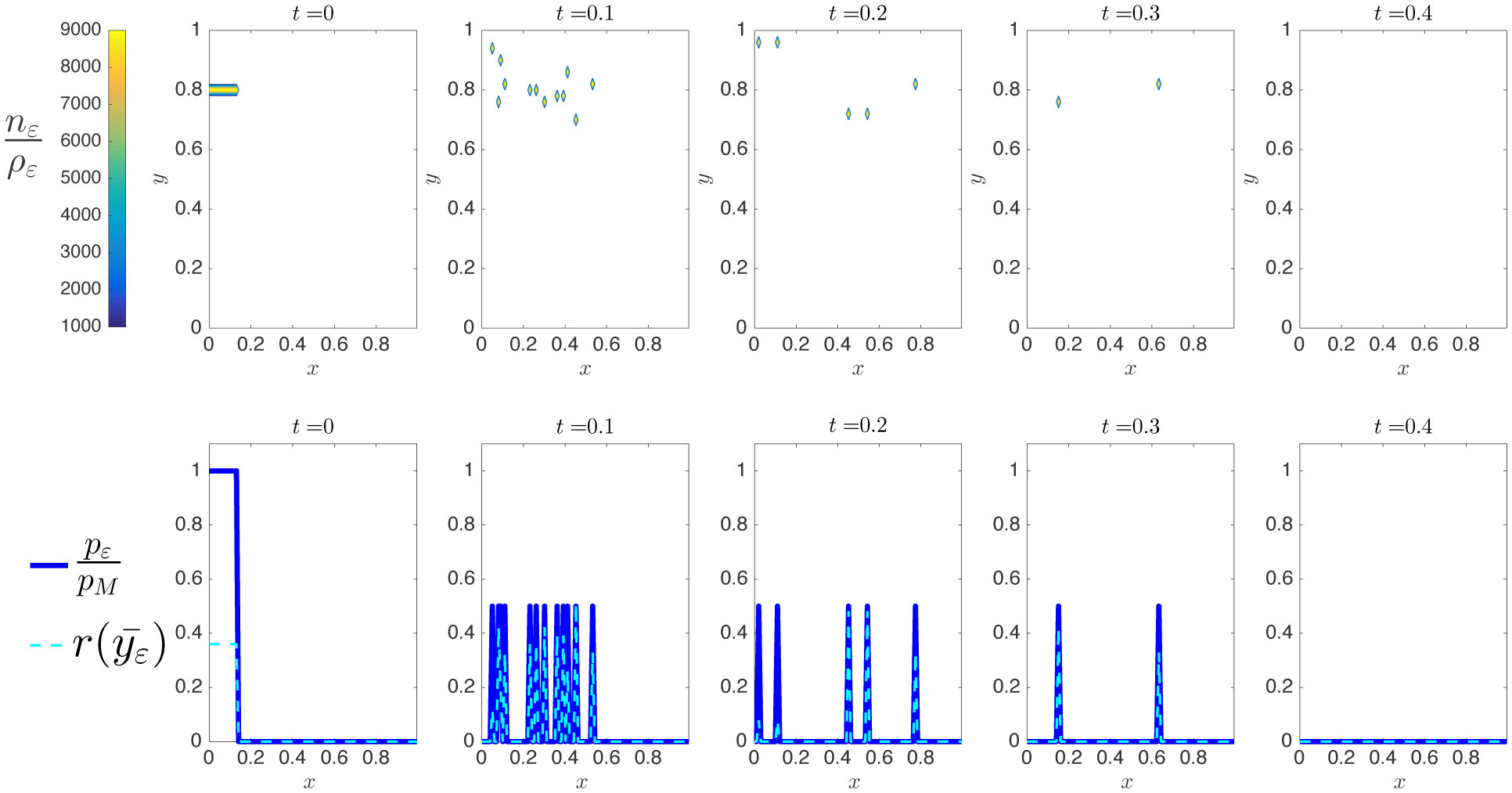
Existing comparative studies between individual-based models of growing cell populations and their continuum counterparts have mainly been focused on homogeneous populations, in which all cells have the same phenotypic characteristics. However, significant intercellular phenotypic variability is commonly observed in cellular systems. In light of these considerations, we develop here an individual-based model for the growth of phenotypically heterogeneous cell populations. In this model, the phenotypic state of each cell is described by a structuring variable that captures intercellular variability in cell proliferation and migration rates. The model tracks the spatial evolutionary dynamics of single cells, which undergo pressure-dependent proliferation, heritable phenotypic changes and directional movement in response to pressure differentials. We formally show that the continuum limit of this model comprises a non-local partial differential equation for the cell population density function, which generalises earlier models of growing cell populations. We report on the results of numerical simulations of the individual-based model which illustrate how proliferation-migration tradeoffs shaping the evolutionary dynamics of single cells can lead to the formation, at the population level, of travelling waves whereby highly-mobile cells locally dominate at the invasive front, while more-proliferative cells are found at the rear. Moreover, we demonstrate that there is an excellent quantitative agreement between these results and the results of numerical simulations and formal travelling-wave analysis of the continuum model, when sufficiently large cell numbers are considered. We also provide numerical evidence of scenarios in which the predictions of the two models may differ due to demographic stochasticity, which cannot be captured by the continuum model. This indicates the importance of integrating individual-based and continuum approaches when modelling the growth of phenotypically heterogeneous cell populations.
Citation: Fiona R Macfarlane, Xinran Ruan, Tommaso Lorenzi. Individual-based and continuum models of phenotypically heterogeneous growing cell populations[J]. AIMS Bioengineering, 2022, 9(1): 68-92. doi: 10.3934/bioeng.2022007
Existing comparative studies between individual-based models of growing cell populations and their continuum counterparts have mainly been focused on homogeneous populations, in which all cells have the same phenotypic characteristics. However, significant intercellular phenotypic variability is commonly observed in cellular systems. In light of these considerations, we develop here an individual-based model for the growth of phenotypically heterogeneous cell populations. In this model, the phenotypic state of each cell is described by a structuring variable that captures intercellular variability in cell proliferation and migration rates. The model tracks the spatial evolutionary dynamics of single cells, which undergo pressure-dependent proliferation, heritable phenotypic changes and directional movement in response to pressure differentials. We formally show that the continuum limit of this model comprises a non-local partial differential equation for the cell population density function, which generalises earlier models of growing cell populations. We report on the results of numerical simulations of the individual-based model which illustrate how proliferation-migration tradeoffs shaping the evolutionary dynamics of single cells can lead to the formation, at the population level, of travelling waves whereby highly-mobile cells locally dominate at the invasive front, while more-proliferative cells are found at the rear. Moreover, we demonstrate that there is an excellent quantitative agreement between these results and the results of numerical simulations and formal travelling-wave analysis of the continuum model, when sufficiently large cell numbers are considered. We also provide numerical evidence of scenarios in which the predictions of the two models may differ due to demographic stochasticity, which cannot be captured by the continuum model. This indicates the importance of integrating individual-based and continuum approaches when modelling the growth of phenotypically heterogeneous cell populations.

| [1] |
Chaplain MAJ, Giverso C, Lorenzi T, et al. (2019) Derivation and application of effective interface conditions for continuum mechanical models of cell invasion through thin membranes. SIAM J Appl Math 79: 2011-2031. https://doi.org/10.1137/19M124263X 
|
| [2] |
Ranft J, Basan M, Elgeti J, et al. (2010) Fluidization of tissues by cell division and apoptosis. Proc Nat Acad Sci USA 107: 20863-20868. https://doi.org/10.1073/pnas.1011086107
|
| [3] |
Giverso C, Ciarletta P (2016) On the morphological stability of multicellular tumour spheroids growing in porous media. Eur Phys J E 39: 92. https://doi.org/10.1140/epje/i2016-16092-7
|
| [4] |
Bubba F, Perthame B, Pouchol C (2020) Hele–shaw limit for a system of two reaction-(cross-) diffusion equations for living tissues equations for living tissues. Arch Ration Mech Anal 236: 735-766. https://doi.org/10.1007/s00205-019-01479-1
|
| [5] |
David N, Ruan X (2022) An asymptotic preserving scheme for a tumor growth model of porous medium type. ESAIM Math Model Numer Anal 56: 121-150. https://doi.org/10.1051/m2an/2021080
|
| [6] |
Lorenzi T, Lorz A, Perthame B (2017) On interfaces between cell populations with different mobilities. Kinet Relat Mod 10: 299-311. https://doi.org/10.3934/krm.2017012
|
| [7] |
Bresch D, Colin T, Grenier E, et al. (2010) Computational modeling of solid tumor growth: the avascular stage. SIAM J Sci Comput 32: 2321-2344. https://doi.org/10.1137/070708895
|
| [8] |
Ciarletta P, Foret L, Ben Amar M (2011) The radial growth phase of malignant melanoma: multi-phase modelling, numerical simulations and linear stability analysis. J R Soc Interface 8: 345-368. https://doi.org/10.1098/rsif.2010.0285
|
| [9] |
Gallinato O, Colin T, Saut O, et al. (2017) Tumor growth model of ductal carcinoma: from in situ phase to stroma invasion. J Theor Biol 429: 253-266. https://doi.org/10.1016/j.jtbi.2017.06.022
|
| [10] | Perthame B (2014) Some mathematical aspects of tumor growth and therapy. ICM 2014-International Congress of Mathematicians . |
| [11] |
Lowengrub JS, Frieboes HB, Jin F, et al. (2009) Nonlinear modelling of cancer: bridging the gap between cells and tumours. Nonlinearity 23: R1. https://doi.org/10.1088/0951-7715/23/1/R01
|
| [12] |
Kuznetsov M, Clairambault J, Volpert V (2021) Improving cancer treatments via dynamical biophysical models. Phys Life Rev 39: 1-48. https://doi.org/10.1016/j.plrev.2021.10.001
|
| [13] |
Lorenzi T (2022) Cancer modelling as fertile ground for new mathematical challenges. Phys Life Rev 40: 3-5. https://doi.org/10.1016/j.plrev.2022.01.003
|
| [14] |
Anderson ARA (2007) A hybrid multiscale model of solid tumour growth and invasion: evolution and the microenvironment. Single-cell-based models in biology and medicine . Switzerland: Birkhäuser Basel 3-28.
|
| [15] |
Van Liedekerke P, Palm MM, Jagiella N, et al. (2015) Simulating tissue mechanics with agent-based models: concepts, perspectives and some novel results. Comput Part Mech 2: 401-444. https://doi.org/10.1007/s40571-015-0082-3
|
| [16] | Inoue M (1991) Derivation of a porous medium equation from many Markovian particles and the propagation of chaos. Hiroshima Math J 21: 85-110. https://doi.org/10.32917/hmj/1206128924 |
| [17] |
Oelschläger K (1989) On the derivation of reaction-diffusion equations as limit dynamics of systems of moderately interacting stochastic processes. Probab Theory Relat Fields 82: 565-586. https://doi.org/10.1007/BF00341284
|
| [18] |
Penington CJ, Hughes BD, Landman KA (2011) Building macroscale models from microscale probabilistic models: a general probabilistic approach for nonlinear diffusion and multispecies phenomena. Phys Rev E 84: 041120. https://doi.org/10.1103/PhysRevE.84.041120
|
| [19] |
Chaplain MAJ, Lorenzi T, Macfarlane FR (2020) Bridging the gap between individual-based and continuum models of growing cell populations. J Math Biol 80: 343-371. https://doi.org/10.1007/s00285-019-01391-y
|
| [20] |
Lorenzi T, Murray PJ, Ptashnyk M (2020) From individual-based mechanical models of multicellular systems to free-boundary problems. Interface Free Bound 22: 205-244. https://doi.org/10.4171/IFB/439
|
| [21] |
Baker RE, Parker A, Simpson MJ (2019) A free boundary model of epithelial dynamics. J Theor Biol 481: 61-74. https://doi.org/10.1016/j.jtbi.2018.12.025
|
| [22] |
Oelschläger K (1990) Large systems of interacting particles and the porous medium equation. J Differ Equ 88: 294-346. https://doi.org/10.1016/0022-0396(90)90101-T
|
| [23] |
Murray PJ, Edwards CM, Tindall MJ, et al. (2009) From a discrete to a continuum model of cell dynamics in one dimension. Phys Rev E 80: 031912. https://doi.org/10.1103/PhysRevE.80.031912
|
| [24] |
Murray PJ, Edwards CM, Tindall MJ, et al. (2012) Classifying general nonlinear force laws in cell-based models via the continuum limit. Phys Rev E 85: 021921. https://doi.org/10.1103/PhysRevE.85.021921
|
| [25] |
Dyson L, Maini PK, Baker RE (2012) Macroscopic limits of individual-based models for motile cell populations with volume exclusion. Phys Rev E 86: 031903. https://doi.org/10.1103/PhysRevE.86.031903
|
| [26] |
Johnston ST, Baker RE, McElwain DL, et al. (2017) Co-operation, competition and crowding: a discrete framework linking Allee kinetics, nonlinear diffusion, shocks and sharp-fronted travelling waves. Sci Rep 7: 1-19. https://doi.org/10.1038/srep42134
|
| [27] |
Johnston ST, Simpson MJ, Baker RE (2012) Mean-field descriptions of collective migration with strong adhesion. Phys Rev E 85: 051922. https://doi.org/10.1103/PhysRevE.85.051922
|
| [28] |
Johnston ST, Simpson MJ, Baker RE (2015) Modelling the movement of interacting cell populations: a moment dynamics approach. J Theor Biol 370: 81-92. https://doi.org/10.1016/j.jtbi.2015.01.025
|
| [29] |
Deroulers C, Aubert M, Badoual M, et al. (2009) Modeling tumor cell migration: from microscopic to macroscopic models. Phys Rev E 79: 031917. https://doi.org/10.1103/PhysRevE.79.031917
|
| [30] |
Drasdo D (2005) Coarse graining in simulated cell populations. Adv Complex Syst 8: 319-363. https://doi.org/10.1142/S0219525905000440
|
| [31] |
Simpson MJ, Merrifield A, Landman KA, et al. (2007) Simulating invasion with cellular automata: Connecting cell-scale and population-scale properties. Phys Rev E 76: 021918. https://doi.org/10.1103/PhysRevE.76.021918
|
| [32] |
Ambrosi D, Preziosi L (2002) On the closure of mass balance models for tumor growth. Math Mod Meth Appl Sci 12: 737-754. https://doi.org/10.1142/S0218202502001878
|
| [33] |
Byrne HM, Chaplain MAJ (1997) Free boundary value problems associated with the growth and development of multicellular spheroids. Eur J Appl Math 8: 639-658. https://doi.org/10.1017/S0956792597003264
|
| [34] |
Byrne HM, Drasdo D (2009) Individual-based and continuum models of growing cell populations: a comparison. J Math Biol 58: 657. https://doi.org/10.1007/s00285-008-0212-0
|
| [35] |
Greenspan HP (1976) On the growth and stability of cell cultures and solid tumors. J Theor Biol 56: 229-242. https://doi.org/10.1016/S0022-5193(76)80054-9
|
| [36] |
Bru A, Albertos S, Subiza JL, et al. (2003) The universal dynamics of tumor growth. Biophys J 85: 2948-2961. https://doi.org/10.1016/S0006-3495(03)74715-8
|
| [37] |
Byrne HM, Preziosi L (2003) Modelling solid tumour growth using the theory of mixtures. Math Med Biol 20: 341-366. https://doi.org/10.1093/imammb/20.4.341
|
| [38] |
Drasdo D, Hoehme S (2012) Modeling the impact of granular embedding media, and pulling versus pushing cells on growing cell clones. New J Phys 14: 055025. https://doi.org/10.1088/1367-2630/14/5/055025
|
| [39] |
Brock A, Chang H, Huang S (2009) Non-genetic heterogeneity—A mutation-independent driving force for the somatic evolution of tumours. Nat Rev Genet 10: 336-342. https://doi.org/10.1038/nrg2556
|
| [40] |
Chisholm RH, Lorenzi T, Clairambault J (2016) Cell population heterogeneity and evolution towards drug resistance in cancer: biological and mathematical assessment, theoretical treatment optimisation. Biochim Biophys Acta Gen Subj 1860: N2627-2645. https://doi.org/10.1016/j.bbagen.2016.06.009
|
| [41] |
Huang S (2013) Genetic and non-genetic instability in tumor progression: Link between the fitness landscape and the epigenetic landscape of cancer cells. Cancer Metastasis Rev 32: 423-448. https://doi.org/10.1007/s10555-013-9435-7
|
| [42] |
Chisholm RH, Lorenzi T, Desvillettes L (2016) Evolutionary dynamics of phenotype-structured populations: From individual-level mechanisms to population-level consequences. Z Angew Math Phys 67: 1-34. https://doi.org/10.1007/s00033-016-0690-7
|
| [43] | Hughes BD (1995) Random walks and random environments: random walks. UK: Oxford University Press. |
| [44] |
Perthame B, Quiros F, Vazquez JL (2014) The Hele–Shaw asymptotics for mechanical models of tumor growth. Arch Ration Mech Anal 212: 93-127. https://doi.org/10.1007/s00205-013-0704-y
|
| [45] |
Basan M, Risler T, Joanny JF, et al. (2009) Homeostatic competition drives tumor growth and metastasis nucleation. HFSP J 3: 265-272. https://doi.org/10.2976/1.3086732
|
| [46] |
Novikov NM, Zolotaryova SY, Gautreau AM (2021) Mutational drivers of cancer cell migration and invasion. Br J Cancer 124: 102-114. https://doi.org/10.1038/s41416-020-01149-0
|
| [47] |
Alfonso JCL, Talkenberger K, Seifert M, et al. (2017) The biology and mathematical modelling of glioma invasion: a review. J R Soc Interface 14: 20170490. https://doi.org/10.1098/rsif.2017.0490
|
| [48] |
Giese A, Bjerkvig R, Berens ME, et al. (2003) Cost of migration: invasion of malignant gliomas and implications for treatment. J Clin Oncol 21: 1624-1636. https://doi.org/10.1200/JCO.2003.05.063
|
| [49] |
Yan M, Yang X, Shen R, et al. (2018) miR-146b promotes cell proliferation and increases chemosensitivity, but attenuates cell migration and invasion via FBXL10 in ovarian cancer. Cell Death Dis 9: 1-17. https://doi.org/10.1038/s41419-018-1093-9
|
| [50] |
Wang SD, Rath P, Lal B, et al. (2012) EphB2 receptor controls proliferation/migration dichotomy of glioblastoma by interacting with focal adhesion kinase. Oncogene 31: 5132-5143. https://doi.org/10.1038/onc.2012.16
|
| [51] |
Godlewski J, Bronisz A, Nowicki MO, et al. (2010) microRNA-451: A conditional switch controlling glioma cell proliferation and migration. Cell Cycle 9: 2814-2820. https://doi.org/10.4161/cc.9.14.12248
|
| [52] |
Aktipis CA, Boddy AM, Gatenby RA, et al. (2013) Life history trade-offs in cancer evolution. Nat Rev Cancer 13: 883. https://doi.org/10.1038/nrc3606
|
| [53] |
Gallaher JA, Brown JS, Anderson ARA (2019) The impact of proliferation-migration tradeoffs on phenotypic evolution in cancer. Sci Rep 9: 1-10. https://doi.org/10.1038/s41598-019-39636-x
|
| [54] |
Gerlee P, Anderson ARA (2009) Evolution of cell motility in an individual-based model of tumour growth. J Theor Biol 259: 67-83. https://doi.org/10.1016/j.jtbi.2009.03.005
|
| [55] | Gerlee Pand Nelander S.The impact of phenotypic switching on glioblastoma growth and invasion. PLoS Comput Biol (2012) 8: e1002556. https://doi.org/10.1371/journal.pcbi.1002556 |
| [56] |
Giese A, Loo MA, Tran N, et al. (1996) Dichotomy of astrocytoma migration and proliferation. Int J Cancer 67: 275-282.
|
| [57] |
Hatzikirou H, Basanta D, Simon M, et al. (2012) ‘Go or Grow’: the key to the emergence of invasion in tumour progression?. Math Med Biol 29: 49-65. https://doi.org/10.1093/imammb/dqq011
|
| [58] |
Orlando PA, Gatenby RA, Brown JS (2013) Tumor evolution in space: The effects of competition colonization tradeoffs on tumor invasion dynamics. Front Oncol 3: 45. https://doi.org/10.3389/fonc.2013.00045
|
| [59] |
Pham K, Chauviere A, Hatzikirou H, et al. (2012) Density-dependent quiescence in glioma invasion: Instability in a simple reaction–diffusion model for the migration/proliferation dichotomy. J Biol Dyn 6: 54-71. https://doi.org/10.1080/17513758.2011.590610
|
| [60] |
Tang M, Vauchelet N, Cheddadi I, et al. (2013) Composite waves for a cell population system modeling tumor growth and invasion. Chinese Ann Math Ser B 34: 295-318. https://doi.org/10.1007/s11401-013-0761-4
|
| [61] |
Ardaseva A, Anderson ARA, Gatenby RA, et al. (2020) Comparative study between discrete and continuum models for the evolution of competing phenotype-structured cell populations in dynamical environments. Phys Rev E 102: 042404. https://doi.org/10.1103/PhysRevE.102.042404
|
| [62] |
Bubba F, Lorenzi T, Macfarlane FR (2020) From a discrete model of chemotaxis with volume-filling to a generalized Patlak–Keller–Segel model. Proc R Soc A 476: 20190871. https://doi.org/10.1098/rspa.2019.0871
|
| [63] |
Macfarlane FR, Chaplain MAJ, Lorenzi T (2020) A hybrid discrete-continuum approach to model Turing pattern formation. Math Biosci Eng 17: 7442-7479. https://doi.org/10.3934/mbe.2020381
|
| [64] |
Stace REA, Stiehl T, Chaplain MAJ, et al. (2020) Discrete and continuum phenotype-structured models for the evolution of cancer cell populations under chemotherapy. Math Mod Nat Phen 15: 14. https://doi.org/10.1051/mmnp/2019027
|
| [65] |
Smith JT, Tomfohr JK, Wells MC, et al. (2004) Measurement of cell migration on surface-bound fibronectin gradients. Langmuir 20: 8279-8286. https://doi.org/10.1021/la0489763
|
| [66] |
Lorenzi T, Perthame B, Ruan X (2021) Invasion fronts and adaptive dynamics in a model for the growth of cell populations with heterogeneous mobility. Eur J Appl Math 2021: 1-18. https://doi.org/10.1017/S0956792521000218
|
| [67] |
Lorenzi T, Painter KJ (2022) Trade-offs between chemotaxis and proliferation shape the phenotypic structuring of invading waves. Int J Non Linear Mech 139: 103885. https://doi.org/10.1016/j.ijnonlinmec.2021.103885
|
| [68] |
Corcoran A, Del Maestro RF (2003) Testing the “go or grow” hypothesis in human medulloblastoma cell lines in two and three dimensions. Neurosurgery 53: 174-185. https://doi.org/10.1227/01.NEU.0000072442.26349.14
|
| [69] |
Hoek KS, Eichhoff OM, Schlegel NC, et al. (2008) In vivo switching of human melanoma cells between proliferative and invasive states. Cancer Res 68: 650-656. https://doi.org/10.1158/0008-5472.CAN-07-2491
|
| [70] |
Stepien TL, Rutter EM, Kuang Y (2018) Traveling waves of a go-or-grow model of glioma growth. SIAM J Appl Math 78: 1778-1801. https://doi.org/10.1137/17M1146257
|
| [71] |
Vittadello ST, McCue SW, Gunasingh G, et al. (2020) Examining go-or-grow using fluorescent cell-cycle indicators and cell-cycle-inhibiting drugs. Biophys J 118: 1243-1247. https://doi.org/10.1016/j.bpj.2020.01.036
|
| [72] | Zhigun A, Surulescu C, Hunt A (2018) A strongly degenerate diffusion-haptotaxis model of tumour invasion under the go-or-grow dichotomy hypothesis. Math Methods Appl Sci 41: 2403-2428. https://doi.org/10.1002/mma.4749 |
| [73] |
Dhruv HD, McDonough Winslow WS, Armstrong B, et al. (2013) Reciprocal activation of transcription factors underlies the dichotomy between proliferation and invasion of glioma cells. PLoS One 8: e72134. https://doi.org/10.1371/journal.pone.0072134
|
| [74] |
Xie Q, Mittal S, Berens ME (2014) Targeting adaptive glioblastoma: an overview of proliferation and invasion. Neuro-Oncol 16: 1575-1584. https://doi.org/10.1093/neuonc/nou147
|
| [75] | Simpson MJ, Baker RE, Buenzli PR, et al. (2022) Reliable and efficient parameter estimation using approximate continuum limit descriptions of stochastic models.. https://doi.org/10.1101/2022.02.02.478913 |
| [76] |
Nardini JT, Baker RE, Simpson MJ, et al. (2021) Learning differential equation models from stochastic agent-based model simulations. J Roy Soc Interface 18: 20200987. https://doi.org/10.1098/rsif.2020.0987
|
| [77] |
Simpson MJ, Sharp JA, Baker RE (2014) Distinguishing between mean-field, moment dynamics and stochastic descriptions of birth–death–movement processes. Phys A Stat Mech Appl 395: 236-246. https://doi.org/10.1016/j.physa.2013.10.026
|
| [78] |
Johnston ST, Simpson MJ, Crampin EJ (2020) Predicting population extinction in lattice-based birth–death–movement models. Proc Roy Soc A 476: 20200089. https://doi.org/10.1098/rspa.2020.0089
|
| [79] |
Powathil GG, Swat M, Chaplain MAJ (2015) Systems oncology: towards patient-specific treatment regimes informed by multiscale mathematical modelling. Sem Cancer Biol 30: 13-20. https://doi.org/10.1016/j.semcancer.2014.02.003
|
| [80] |
Rejniak KA, Anderson ARA (2011) Hybrid models of tumor growth. Wiley Interdiscip Rev Syst Biol Med 3: 115-125. https://doi.org/10.1002/wsbm.102
|
| [81] |
Jafari Nivlouei S, Soltani M, Carvalho J, et al. (2021) Multiscale modeling of tumor growth and angiogenesis: Evaluation of tumor-targeted therapy. PLoS Comp Biol 17: e1009081. https://doi.org/10.1371/journal.pcbi.1009081
|
| [82] | Jafari Nivlouei S, Soltani M, Shirani E, et al. (2022) A multiscale cell-based model of tumor growth for chemotherapy assessment and tumor-targeted therapy through a 3D computational approach. Cell Prolif 2022: e13187. https://doi.org/10.1111/cpr.13187 |
 bioeng-09-01-007-s001.pdf bioeng-09-01-007-s001.pdf |
 |
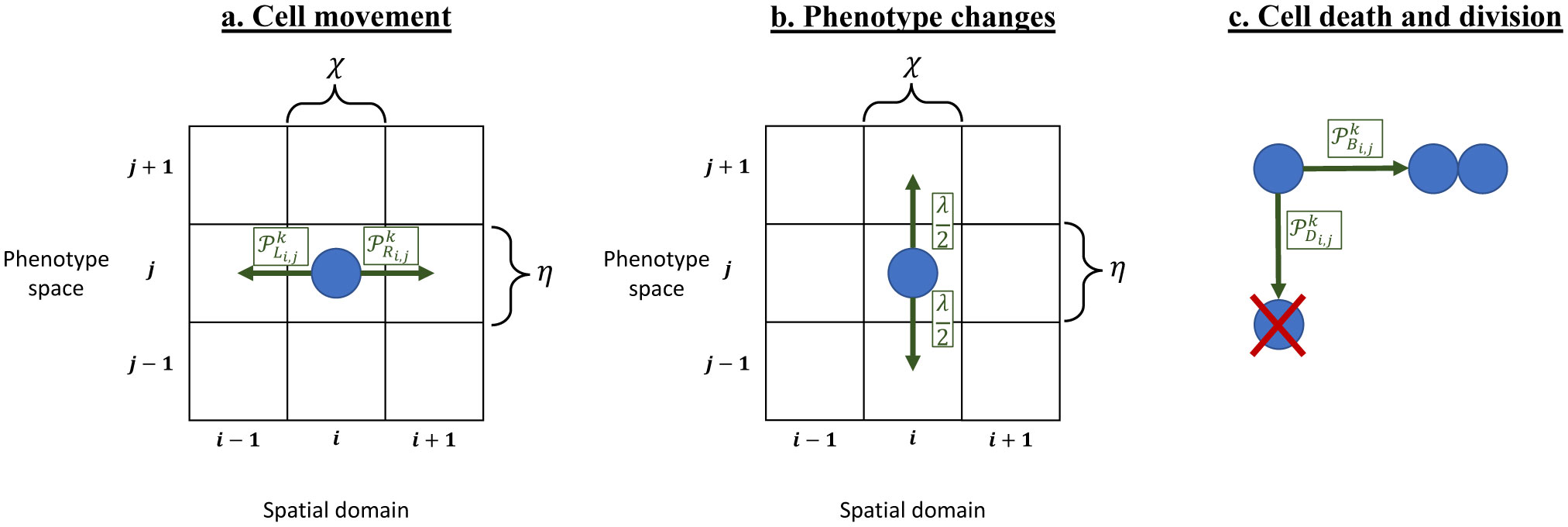
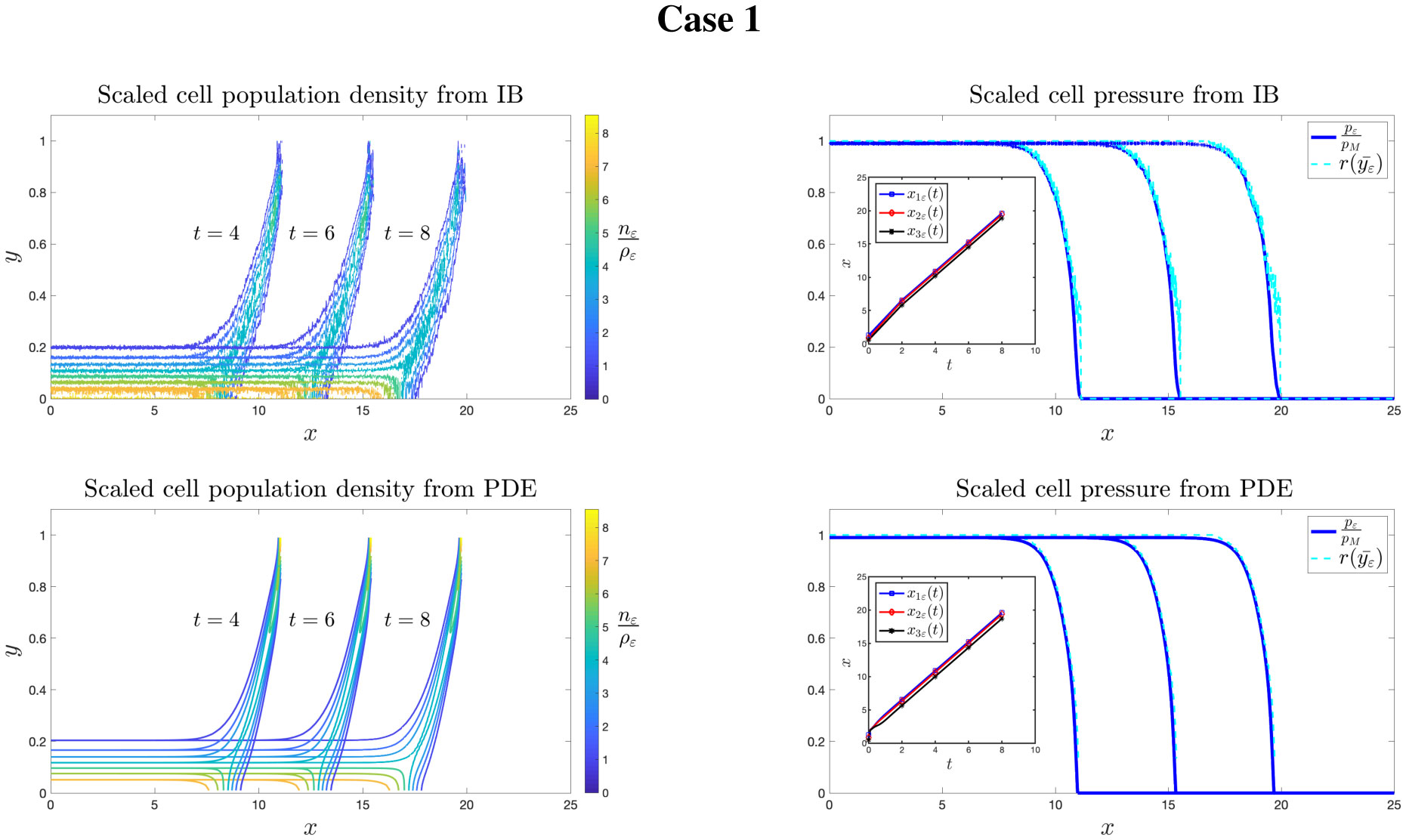
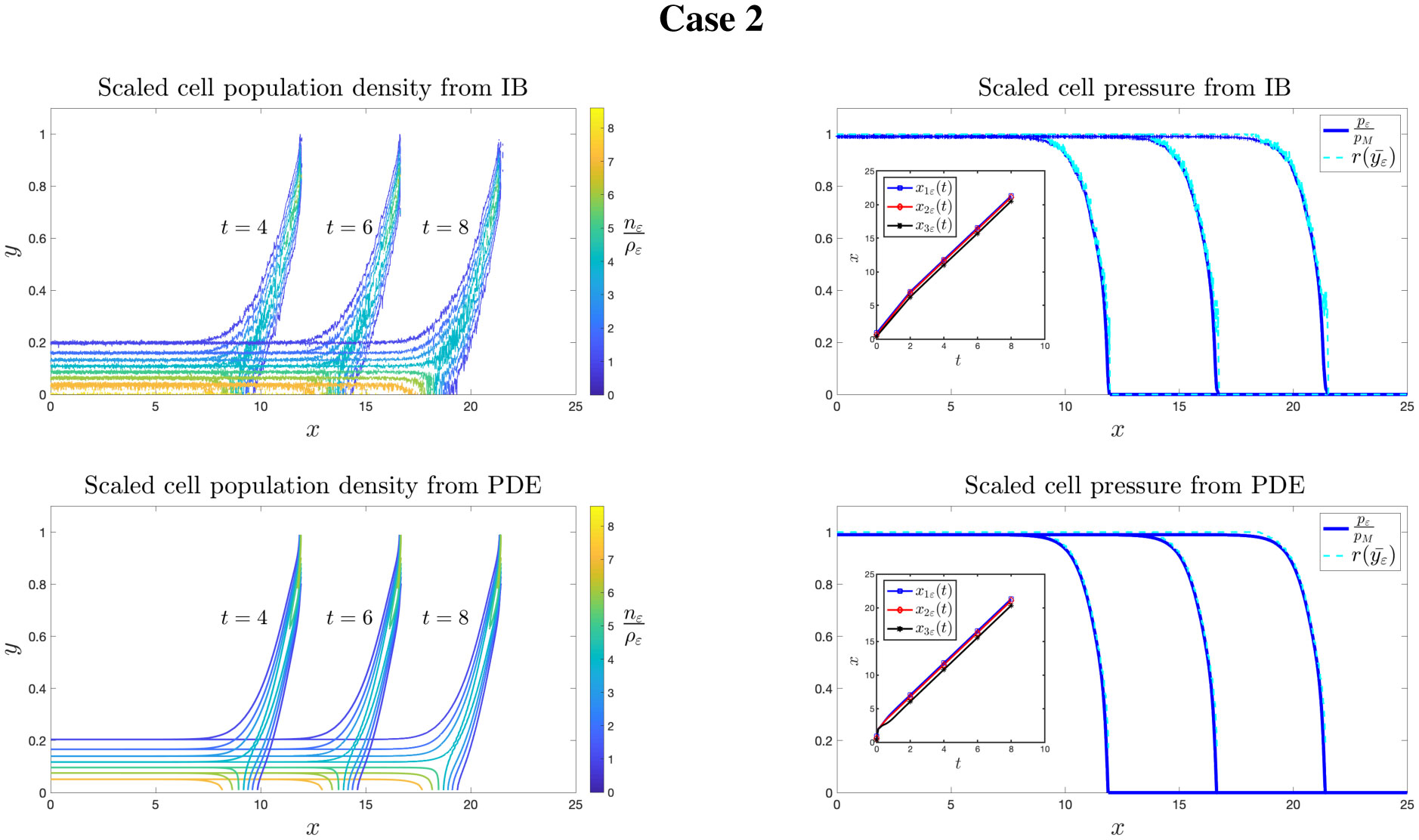
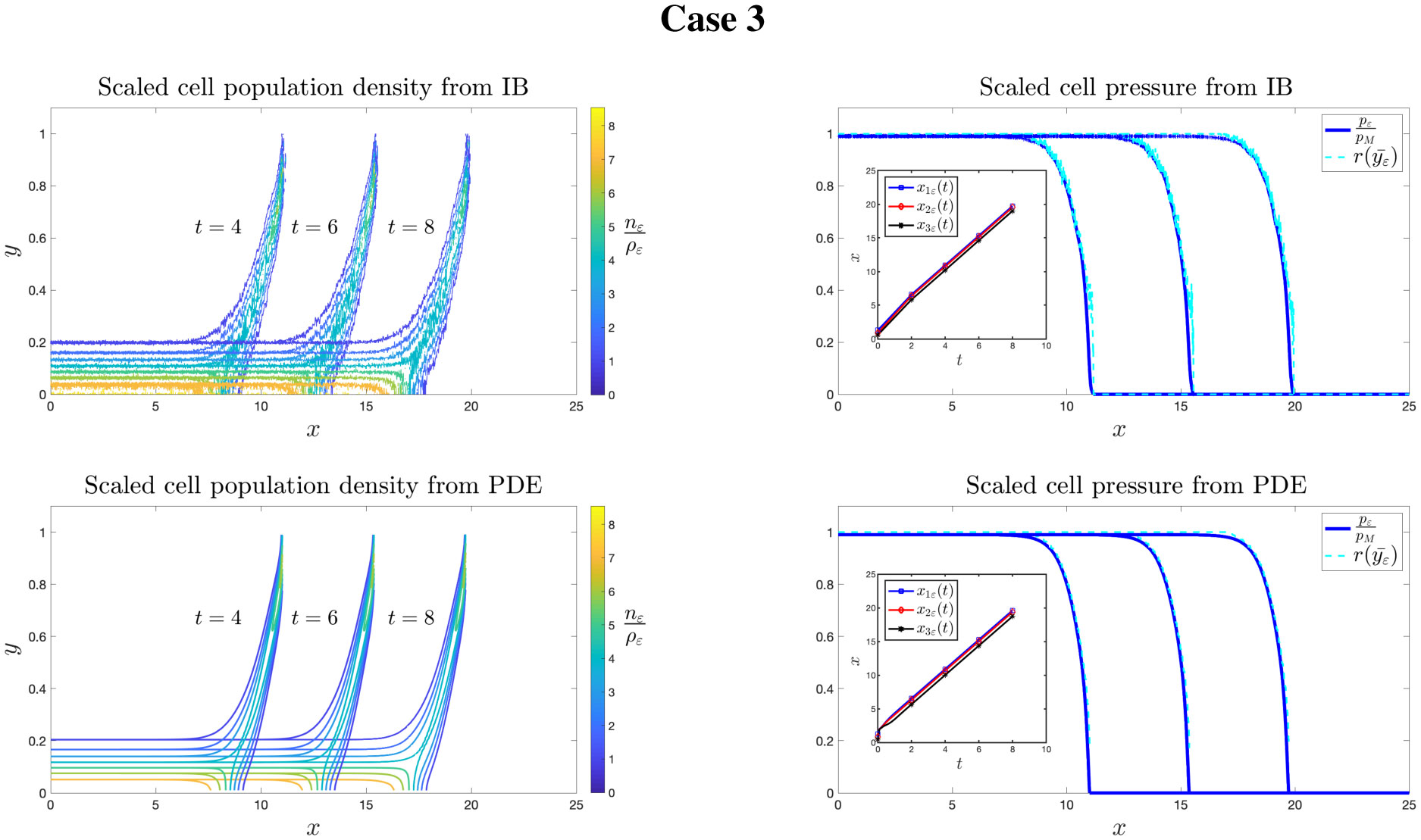
Figures(8)

Fiona R Macfarlane, Xinran Ruan, Tommaso Lorenzi. Individual-based and continuum models of phenotypically heterogeneous growing cell populations[J]. AIMS Bioengineering, 2022, 9(1): 68-92. doi: 10.3934/bioeng.2022007







 DownLoad:
DownLoad: