Citation: Madhuri Sharon, Farha Modi, Maheshwar Sharon. Titania based nanocomposites as a photocatalyst: A review[J]. AIMS Materials Science, 2016, 3(3): 1236-1254. doi: 10.3934/matersci.2016.3.1236

| [1] |
Hashimoto K, Irie H, Fujishima A (2005) TiO2 photocatalysis: An historical overview and future prospects. Jpn J Appl Phys 44: 8269–8285. doi: 10.1143/JJAP.44.8269

|
| [2] |
Chen X, Mao SS (2007) Titanium dioxide nanomaterials: synthesis, properties, modifications, and applications. Chem Rev 107: 2891–959. doi: 10.1021/cr0500535
|
| [3] |
Fox MA, Dulay MT (1993) Heterogeneous photocatalysis. Chem Rev 93: 341–357. doi: 10.1021/cr00017a016
|
| [4] |
Hoffmann MR, Martin ST, Choi W, et al. (1995) Environmental applications of semiconductor photocatalysis. Chem Rev 95: 69–96. doi: 10.1021/cr00033a004
|
| [5] |
Lee Y, Misook K (2010) The optical properties of nanoporous structured Titanium dioxide and the photovoltaic efficiency on DSSC. Mater Chem Phys 122: 284–289. doi: 10.1016/j.matchemphys.2010.02.050
|
| [6] |
Fujishima A, Honda K (1972) Electrochemical photolysis of water at a semiconductor electrode. Nature 238: 37–38. doi: 10.1038/238037a0
|
| [7] |
Gabor A, Somorjai A, Contreras M, et al. (2006) Clusters, surfaces, and catalysis. P Natl Acad Sci USA 103: 10577–10583. doi: 10.1073/pnas.0507691103
|
| [8] |
Mills A, Hunte SL (1997) An overview of semiconductor photocatalysis. J Photoch Photobiol A 108: 1–35. doi: 10.1016/S1010-6030(97)00118-4
|
| [9] |
Burda C, Chen X, Narayanan R, et al. (2005) Chemistry and properties of nanocrystals of different shapes. Chem Rev 105: 1025–1102. doi: 10.1021/cr030063a
|
| [10] |
Pelizzetti E, Minero C (1994) Metal oxides as photocatalysts for environmental detoxification. Comment Inorg Chem 15: 297–337. doi: 10.1080/02603599408035846
|
| [11] |
Hisatomi T, Kubota J, Domen K (2014) Recent advances in semiconductors for photocatalytic and photoelectrochemical water splitting. Chem Soc Rev 43: 7520–7535. doi: 10.1039/C3CS60378D
|
| [12] | Ramírez H, Ramírez M (2015) Photocatalytic Semiconductors: Synthesis, Characterization, and Environmental Applications. Springer International Publishing, ISBN 978-3-319-10999-2. |
| [13] |
Chen H, Nanayakkara CE, Grassian VH (2012) Titanium dioxide photocatalysis in atmospheric chemistry. Chem Rev 112: 5919–5948. doi: 10.1021/cr3002092
|
| [14] |
Pelaez M, Nolan NT, Pillai SC, et al. (2012) A review on the visible light active Titanium dioxide photocatalysts for environmental applications. Appl Catal B 125: 331–349. doi: 10.1016/j.apcatb.2012.05.036
|
| [15] |
Kalathil S, Khan MM, Ansari SA, et al. (2013) Band gap narrowing of Titanium dioxide (TiO2) nanocrystals by electrochemically active biofilm and their visible light activity. Nanoscale 5: 6323–6326. doi: 10.1039/c3nr01280h
|
| [16] |
Khan MM, Ansari SA, Pradhan D, et al. (2014) Band gap engineered TiO2 nanoparticles for visible light induced photoelectrochemical and photocatalytic studies. J Mater Chem A 2: 637–644. doi: 10.1039/C3TA14052K
|
| [17] |
Carp O, Huisman CL, Reller A (2004) Photoinduced reactivity of Titanium dioxide. Prog Solid State Ch 32: 33–177. doi: 10.1016/j.progsolidstchem.2004.08.001
|
| [18] | Chen Q, Peng LM (2007) Structure and applications of titanate and related nanostructures. Int J Nanotechnol 4: 261–270. |
| [19] | Amaratunga P (2010) Synthesis and characterization of monolayer protected gold nanoparticles and a Gold-Titanium dioxide nanocomposite intended for photovoltaic degradation of environmental pollutants. Arch Microbiol 151: 77–83. |
| [20] |
Jang JS, Sun S, Choi H, et al. (2006) A composite deposit photocatalyst of CdS nanoparticles deposited on TiO2 Nanosheets. J Nanosci Nanotechno 6: 3642–3646. doi: 10.1166/jnn.2006.073
|
| [21] | Inumaru K, Kasahara T, Yasui M, et al. (2005) Direct nanocomposite of crystallite TiO2 particles and mesoporous silica as a molecular selective and highly reactive photocatalyst. Chem Commun 2005: 2132–1233. |
| [22] | Pradhan S, Ghosh D, Chen S (2009) Janus nanostructures based on Au-TiO2 heterodimers and their photocatalytic activity in the oxidation of methanol. ACS Appl Mater Inter 1: 2060–2065. |
| [23] | Fujishima A, Rao TN, Tryk DA (2000) Titanium dioxide photocatalysis. J Photoch Photobio C 1: 1–21. |
| [24] |
Wang S, Zhou S (2011) Photodegradation of Methyl orange by photocatalyst of CNTs/P-TiO2 under UV and visible-light irradiation. J Hazard Mater 185: 77–85. doi: 10.1016/j.jhazmat.2010.08.125
|
| [25] | Ibrahim SA, Sreekantan S (2010) Effect of pH on TiO2 nanoparticles via sol-gel method. Adv Mater Res 173: 184–189. |
| [26] |
Niederberger M, Bartl MH, Stucky GD (2002) Benzyl alcohol and transition metal chlorides as a versatile reaction system for the nonaqueous and low-temperature synthesis of crystalline nano-objects with controlled dimensionality. J Am Chem Soc 124: 13642–13643. doi: 10.1021/ja027115i
|
| [27] |
Parala H, Devi A, Bhakta R, et al. (2002) Synthesis of nano-scale TiO2 particles by a non-hydrolytic approach. J Mater Chem 12: 1625–1627. doi: 10.1039/b202767d
|
| [28] |
Lei H, Hou Y, Zhu M, et al. (2005) Formation and transformation of ZnTiO3 prepared by sol-gel process. Mater Lett 59: 197–200. doi: 10.1016/j.matlet.2004.07.046
|
| [29] |
Arnal P, Corriu RJP, Leclercq D, et al. (1996) Preparation of anatase, brookite and rutile at low temperature by non-hydrolytic sol-gel methods. J Mater Chem 6: 1925–1932. doi: 10.1039/JM9960601925
|
| [30] | Arnal P, Corriu RJP, Leclercq D, et al. (1997) A solution chemistry study of nonhydrolytic Sol-Gel routes to Titania.Chem Mater9: 694–698. |
| [31] |
Hay JN, Raval HM (1998) Preparation of inorganic oxides via a non-hydrolytic sol-gel route. J Sol-Gel Sci Techn 13: 109–112. doi: 10.1023/A:1008615708489
|
| [32] |
Hay JN, Raval HM (2001) Synthesis of organic-inorganic hybrids via the non-hydrolytic sol-gel process. Chem Mater 13: 3396–3403. doi: 10.1021/cm011024n
|
| [33] | Lafond V, Mutin PH, Vioux A (2002) Non-hydrolytic sol-gel routes based on alkyl halide elimination: Toward better mixed oxide catalysts and new supports—Application to the preparation of a SiO2-TiO2 epoxidation catalyst. J Mol Cata A-Chem 182: 81–88. |
| [34] |
Trentler TJ, Denler TE, Bertone JF, et al. (1999) Synthesis of TiO2 nanocrystals by nonhydrolytic solution-based reactions. J Am Chem Soc 121: 1613–1614. doi: 10.1021/ja983361b
|
| [35] |
Byrappa K, Adschiri T (2007) Hydrothermal technology for nanotechnology. Prog Cryst Growth Ch 53: 117–166. doi: 10.1016/j.pcrysgrow.2007.04.001
|
| [36] |
Andersson M, Österlund L, Ljungström S, et al. (2002) Preparation of nanosize anatase and rutile TiO2 by hydrothermal treatment of microemulsions and their activity for photocatalytic wet oxidation of phenol. J Phys Chem B 106: 10674–10679. doi: 10.1021/jp025715y
|
| [37] |
Yong CS, Park MK, Lee SK, et al. (2003) Preparation of size-controlled TiO2 nanoparticles and derivation of optically transparent photocatalytic films. Chem Mater 15: 3326–3331. doi: 10.1021/cm030171d
|
| [38] |
Cot F, Larbot A, Nabias G (1998) Preparation and characterization of colloidal solution derived crytalline titania powder. J Euro Ceram Soc 18: 2175–2181. doi: 10.1016/S0955-2219(98)00143-5
|
| [39] |
Yang J, Mei S, Ferreira JMF (2000) Hydrothermal synthesis of nanosized titania powders: influence of peptization and peptizing agents on the crystalline phases and phase transitions. J Am Ceram Soc 83: 1361–1268. doi: 10.1111/j.1151-2916.2000.tb01394.x
|
| [40] | Yang J, Mei S, Ferreira JMF (2001) Hydrothermal synthesis of nanosized titania powders: Influence of tetraalkyl ammonium hydroxide on particle characteristics. J Am Ceram Soc 84: 1696–1702. |
| [41] |
Yang J, Di L (2002) Rapid synthesis of nanocrystalline TiO2/SnO2 binary oxide and their photoinduced decompositopn of methyl orange. J Solid State Chem 165: 193–198. doi: 10.1006/jssc.2001.9526
|
| [42] | Yang TY, Lin HM, Wei BY, et al. (2003) UV enhancement of the gas sensing properties of nano-TiO2. Rev Adv Mater Sci 4: 48–54. |
| [43] | Liveri VT (2002) Reversed micelles as nanometer-size solvent media. In Nano-Surface Chemistry. Rosoff M, Ed. Marcel Dekker: New York, 473–385. |
| [44] |
Zhang D, Limin Q, Jiming M, et al. (2002) Formation of crystalline nanosized titania in reverse micelles at room temperature. J Mater Chem 12: 3677–3680. doi: 10.1039/b206996b
|
| [45] | Hong SS, LeeSL, Lee GD (2003) Photocatalytic degradation of p-Nitrophenol over Titanium dioxide prepared by reverse microemulsion method using non-ionic suefactant with different hydrophpsilic groups. React Kinet Cat Lett 80: 145–151. |
| [46] |
Kim KD, Kim TH (2005) Comparison of the growth mechanism of TiO2-coated SiO2 particles prepared by Sol-gel process and water-in-oil type microemulsion method. Colloid Surface A 255: 131–137. doi: 10.1016/j.colsurfa.2004.12.036
|
| [47] | Li GL, Wang GH (1999) Synthesis of nanometer-sized TiO2 particles by a microemulsion method. Nanostruct Mater 11: 663–668. |
| [48] | Li Y, Cureton LT, Sun YP (2004) Improving photoreduction of CO2 with homogeneously dispersed nanoscale TiO2 catalysts. Chem Commun 2004: 1234–1235. |
| [49] | Chen X, Mao SS (2007) Titanium dioxide nanomaterials:? Synthesis, properties modifications, and applications. Chem Rev 107: 2891–2959. |
| [50] |
Lim KT, Ha SH (2004) Synthesis of TiO2 nanoparticles utilizing hydrated reverse micelles in CO2. Langmuir 20: 2466–2471. doi: 10.1021/la035646u
|
| [51] |
Yu JC, Zhang L, Yu J (2002) Direct sonochemical preparation and characterization of highly active mesoporous TiO2 with a bicrystalline framework. Chem Mater 14: 4647–4653. doi: 10.1021/cm0203924
|
| [52] |
Li XL, Peng Q, Yi JX, et al. (2006) Near monodisperse TiO2 nanoparticles and nanorods. Chem A Euro J 12: 2111–2395. doi: 10.1002/chem.200690023
|
| [53] |
Xu J, Ao Y, Fu D, et al. (2008) Synthesis of fluorinedoped titania-coated activated carbon under low temperature with high photocatalytic activity under visible light. J Phys Chem Sol 69: 2366–2370. doi: 10.1016/j.jpcs.2008.03.017
|
| [54] |
Wang X, Zhuang J, Peng Q, et al. (2005) A general strategy for nanocrystal synthesis. Nature 437: 121–124. doi: 10.1038/nature03968
|
| [55] | Krishna KM, Paii VA, Marathe VR, et al. (1990) Atheoretical approach to design of reduced band gap non corrosive electrode for photoelectrochemical solar cell. Int J Quantum Chem 24: 419–427. |
| [56] | Sharon M, Krishna KM, Mishra MK, et al. (1992) Theoretical investigation of optimal mixing ratio for PbO2 and TiO2 to produce a low band gap noncorrosive photoelectrode. J Chem Phys 163: 401–412. |
| [57] |
Krishna KM, Sharon M, Mishra MK (1995) Preparation and characterization of a PbTiO3 + PbO mixed oxide photoelectrode. J Electroanalytic Chem 391: 93–99. doi: 10.1016/0022-0728(95)03905-V
|
| [58] |
Sharon M, Krishna KM, Mishra MK (1996) Preparation and characterization of mixed oxides obtained from various molar mixtures of beta-PbO2 and TiO2. J Phys Chem Solids 57: 615–626. doi: 10.1016/0022-3697(95)00272-3
|
| [59] | Sharon M, Krishna KM, Mishra MK (1996) Pb1?xTixO: a new photoactive phase. J Mater Sci Lett 15: 1084–1087. |
| [60] |
Wei XX, Cui H, Guo S, et al. (2013) Hybrid BiOBr-TiO2 nanocomposites with high visible lightphotocatalytic activity for water treatment. J Hazard Mater 263: 650–658. doi: 10.1016/j.jhazmat.2013.10.027
|
| [61] |
Chakraborty AK, Hossain ME, Rhaman MM, et al. (2014) Fabrication of Bi2O3/TiO2 nanocomposites and their applications to the degradation of pollutants in air and water under visible-light. J Environ Sci 26: 458–465. doi: 10.1016/S1001-0742(13)60428-3
|
| [62] | Khan B, Ashraf U (2015) Sol-gel synthesis and characterization of nanocomposites of Cu/TiO2 and Bi/TiO2 metal oxides as photocatalysts. Int J Sci Technol 4: 40–48. |
| [63] | Dresselhaus MS, Dresselhaus G (2001) Carbon nanotubes: Synthesis, Structure, Properties and Applications: Topics in Applied Physics, Springer-Verlag. ISBN 3-54041-086-4, Berlin. |
| [64] |
Saleh TA, Gupta VK (2011) Functionalization of tungsten oxide into MWCNT and its application for sunlight-induced degradation of rhodamine B. J Colloid Interface Sci 362: 337–344. doi: 10.1016/j.jcis.2011.06.081
|
| [65] |
Yu JC, Zhang L, Zheng Z, et al. (2003) Synthesis and characterization of phosphate mesoporous Titanium dioxide with high photocatalytic activity. Chem Mater 15: 2280–2286. doi: 10.1021/cm0340781
|
| [66] | Lin L, Lin W, Zhu YX, et al. (2005)Phosphor-doped titania—a novel photocatalyst active in visible light. Chem Lett 34: 284–285. |
| [67] |
Korosi L, Oszko A, Galbacs G, et al. (2007) Structural properties and photocatalytic behavior of phosphate-modified nanocrystalline titania films. Appl Catal B 77: 175–183. doi: 10.1016/j.apcatb.2007.07.019
|
| [68] |
Lin L, Lin W, Xie JL, et al. (2007) Photocatalytic properties of phosphor-doped titania nanoparticles. Appl Catal B 75: 52–58. doi: 10.1016/j.apcatb.2007.03.016
|
| [69] |
Jin C, Zheng RY, Guo Y, et al. (2009) Hydrothermal synthesis and characterization of phosphorous-doped TiO2 with high photocatalytic activity for methylene blue degradation. J Mol Catal A 313: 44–48. doi: 10.1016/j.molcata.2009.07.021
|
| [70] |
Wang S, Zhou S (2011) Photodegradation of methyl orange by photocatalyst of CNTs/P-TiO2 under UV and visible-light irradiation. J Hazard Mater 185: 77–85. doi: 10.1016/j.jhazmat.2010.08.125
|
| [71] | Sharon M, Datta S, Shah S, et al. (2007) Photocatalytic degradation of E. coli and S. aureus by multi walled carbon nanotubes. Carbon Letts 8: 184–190. |
| [72] | Oza G, Pandey S, Gupta A, et al. (2013) Photocatalysis-assisted water filtration: Using TiO2-coated vertically aligned multi-walled carbon nanotube array for removal of Escherichia coli O157:H7. Mater Sci Eng C-Mater 33: 4392–4400. |
| [73] | Cong Y, Li X, Qin Y, et al. (2011) Carbon-doped TiO2 coating on multiwalled carbon nanotubes with higher visible light photocatalytic activity. Appl Catal B-Environ 107: 128–134. |
| [74] | Mamba G, Mbianda XY, Mishra AK (2014) Gadolinium nanoparticles decorated multiwalled carbon nanotube/titania nanocomposite for degradation of methylene blue in water under simulated solar light. Environ Sci Pollut Res 21: 5597–5609. |
| [75] |
Mamba G, Mbianda XY, Mishra AK (2015) Photocatalytic degradation of diazo dye naphthol blue black in water using MWCNT/Gd, N, S-TiO2 nanocomposite under simulated solar light. J Environ Sci 33: 219–228. doi: 10.1016/j.jes.2014.06.052
|
| [76] |
Czech B, Buda W (2015) Photocatalytic treatment of pharmaceutical wastewater using new multiwall-carbon nanotubes/TiO2/SiO2 nanocomposite. Environ Res 137: 176–184. doi: 10.1016/j.envres.2014.12.006
|
| [77] | Ptrovic M, Radjenovic J, Postigo C, et al. (2008) Emerging contaminants in waste waters: sources and occurrence. In: Barcello D, Ptrovic M, Eds. Emerging contaminants from Industrial and Municipal Waste. Springer, Berlin, Heidelberg, 1–35. |
| [78] |
Gadipelly C, Perez-Gonzalez A, Yadav GD, et al. (2014) Pharmaceutical industry waste water—reviews of the technology for water treatment and re-use. Ind Eng Chem Res 53: 11571–11592. doi: 10.1021/ie501210j
|
| [79] | Krishamoorthy K, Mohan R, Kim SJ (2001) Graphene oxide as photocatalytic material. Appl Phys Lett 98: 244101–114312. |
| [80] | Stengl V, Bakardjieva S, Gryger TM, et al. (2013) TiO2-graphene oxide nanocompositeas advanced photocatalytic materials. Chem Central J 7: 41–53. |
| [81] |
Zhang Y, Zhou Z, Chen T, et al. (2014) Graphene TiO2 nanocomposite with high photocatalytic activity for degradation of sodium pentachlorophenol. J Environ Sci 26: 2114–2122. doi: 10.1016/j.jes.2014.08.011
|
| [82] |
Stein A (2003) Advances in microporous and mesoporous solids—Highlights of recent progress. Adv Mater 15: 763–775. doi: 10.1002/adma.200300007
|
| [83] | Stein A, Melde BJ, Schroden RC (2003) Hybrid inorganic-organic mesoporous silicates—nanoscopic reactors coming of age. Adv Mater 12: 1403–1419. |
| [84] | Inumaru K, Kasahara T, Yasui M, et al. (2005) Direct nanocomposite of crystallite TiO2 particles and mesoporous silica as a molecular selective and highly active photocatalyst. Chem Commun 2005: 2131–2133. |
| [85] | Mohseni A, Malekina L, Fazaeli R, et al. (2013) Synthesis TiO2/SiO2/Ag nanocomposite by sonochemical method and investigation of photo-catalyst effect in waste water treatment. Nanocon 10: 16–18. |
| [86] | Li K, Huang C (2000) Selective oxidation of Hydrogen Sulfide to sulphur over LaVO4 catalyst: Promotional effect of Antimony oxide addition. Ind Eng Chem Res 45: 7096–7100. |
| [87] | Ye JH, Zhou ZG, Oshikiri M, et al. (2003) New visible light driven semiconductor photocatalyst and their application as functional eco-material. Mater Sci Forum 423: 825–830. |
| [88] |
Huang H, Li D, Lin Q, et al. (2009) Efficient degradation of Benzene over LaVO4/TiO2 nano-crystalline heterojunction photocatalyst under visible light irradiation. Envron Sci Technol 43: 4164–4168. doi: 10.1021/es900393h
|
| [89] | Visa M, Duta A (2013) Methyl orange and Cadmium simultaneous removal using fly ash and Photo-Fenton system. J Hazard Mater 244–245: 773–779. |
| [90] |
Visa M (2012) Tailoring fly ash activated with bentonite as adsorbent for complex waste water treatment. Appl Surf Sci 263: 753–762. doi: 10.1016/j.apsusc.2012.09.156
|
| [91] | Visa M, Andronic L, Duta A (2015) Fly ash-TiO2 nanocomposite material for multi-pollutants water treatment. J Environ Manage 150: 336–343. |
| [92] |
Kaplan R, Erjavec B, Drazic G, et al. (2016) Simple synthesis of Anatase/rutile/brookite TiO2 nanocomposite with superior mineralization potential for photocatalytic degradation of water pollutants. Appl Catal B-Environ 181: 465–474. doi: 10.1016/j.apcatb.2015.08.027
|
| [93] | Yu J, Qi L (2009) Template free fabrication of hierarchically flower like tungsten tri oxide assemblies with enhanced visible-light-driven photocatalytic activity. J Hazard Mater 169: 221–227. |
| [94] |
Vicaksana Y, Liu S, Scott J, et al. (2014) Tungsten trioxide as a visible light photocatalyst for volatile organic carbon removal. Molecules 19: 17747–17762. doi: 10.3390/molecules191117747
|
| [95] | Sajjad AKL, Sajjad S, Tian B, et al. (2010) Comparative studies of operational parameters of degradation of azo-dyes in visible light by highly efficient WOx/TiO2 photocatalyst. J Hazard Mater 177: 781–791. |
| [96] |
Zhao G, Jr SES (1998) Multiple parameters for the comprehensive evaluation of the susceptibility of Escherichia coli to the silver ion. Biometals 11: 27–32. doi: 10.1023/A:1009253223055
|
| [97] |
Yamanaka M, Hara K, Kudo J (2005) Bactericidal actions of a Silver ion solution on Escherichia coli, studied by Energy-Filtering Transmission Electron Microscopy and Proteomic Analysis. Appl Environ Microb 71: 7589–7593. doi: 10.1128/AEM.71.11.7589-7593.2005
|
| [98] |
Jung WK, Koo HC, Kim KW, et al. (2008) Antibacterial activity and mechanism of action of the silver ion in Staphylococcus aureus and Escherichia coli. Appl Environ Microb 74: 2171–2178. doi: 10.1128/AEM.02001-07
|
| [99] | Liu SX, Qu ZP, Han WX, et al. (2004) A mechanism for enhanced photocatalytic activity of silver loaded titania dioxide. Catal Today 93–95: 877–884. |
| [100] |
Akhavan O (2009) Lasting antibacterial activities of Ag-TiO2/Ag/a-TiO2 nanocomposite thin film photocatalysts under solar light irradiation. J Colloid Interf Sci 336: 117–124. doi: 10.1016/j.jcis.2009.03.018
|
| [101] | Xiang Q, Yu J, Cheng B, et al. (2010) Microwave hydrothermal preparation of Visible-light photocatalytic activity of Ag-TiO2 nanocomposite hollow sphere. Chem Asian J 5: 1466–1474. |
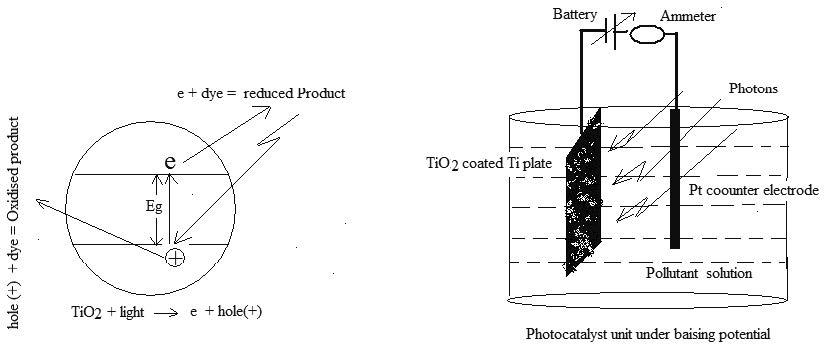
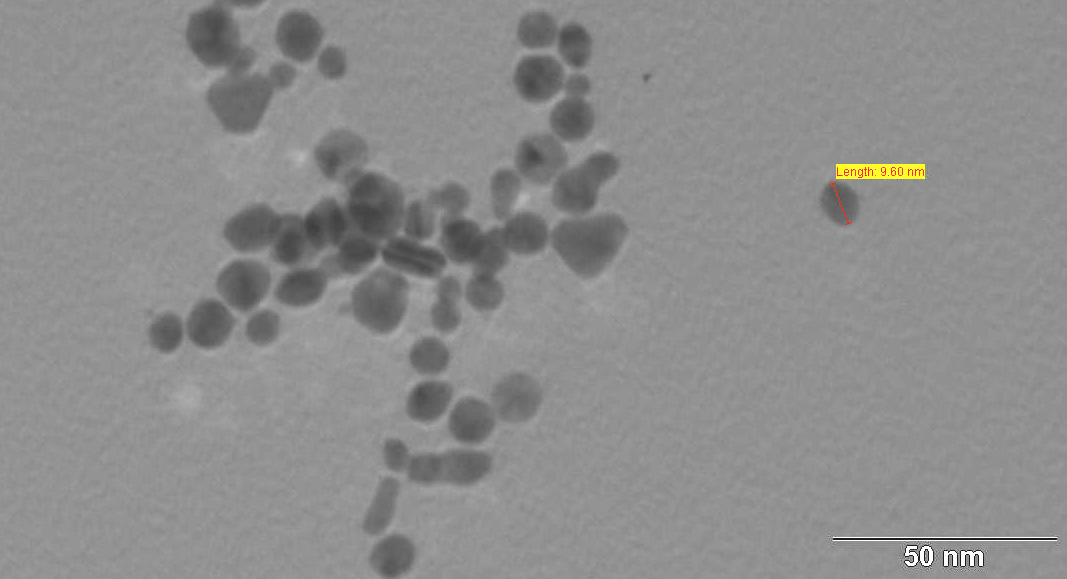
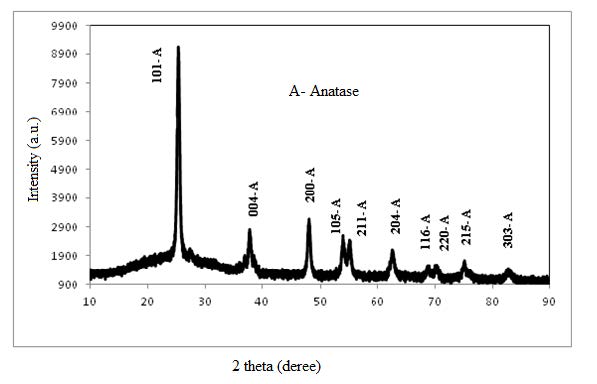
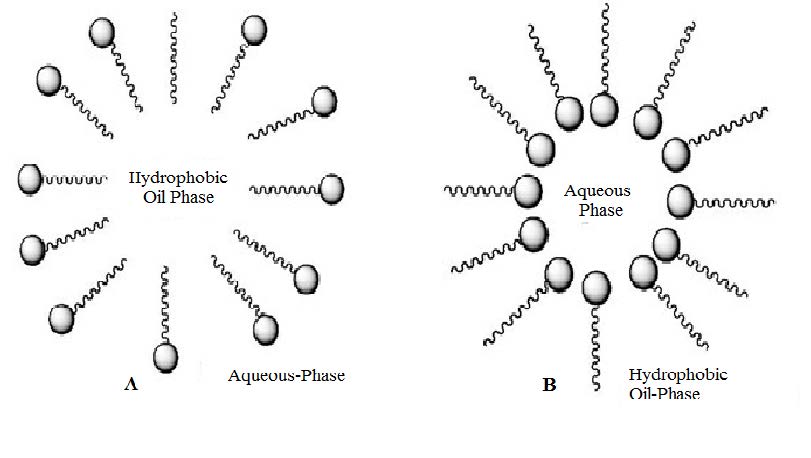
Figures(5)

Madhuri Sharon, Farha Modi, Maheshwar Sharon. Titania based nanocomposites as a photocatalyst: A review[J]. AIMS Materials Science, 2016, 3(3): 1236-1254. doi: 10.3934/matersci.2016.3.1236







 DownLoad:
DownLoad: