Citation: Sarmed H. Kathem, Ashraf M. Mohieldin, Surya M. Nauli. The Roles of Primary cilia in Polycystic Kidney Disease[J]. AIMS Molecular Science, 2014, 1(1): 27-46. doi: 10.3934/molsci.2013.1.27

| [1] |
Lentine KL, Xiao H, Machnicki G, et al. (2010) Renal function and healthcare costs in patients with polycystic kidney disease. Clin J Am Soc Nephrol 5: 1471-1479. doi: 10.2215/CJN.00780110

|
| [2] |
Torres VE, Harris PC (2009) Autosomal dominant polycystic kidney disease: the last 3 years. Kidney Int 76: 149-168. doi: 10.1038/ki.2009.128
|
| [3] |
Grantham JJ, Torres VE, Chapman AB, et al. (2006) Volume progression in polycystic kidney disease. N Engl J Med 354: 2122-2130. doi: 10.1056/NEJMoa054341
|
| [4] |
Churchill DN, Bear JC, Morgan J, et al. (1984) Prognosis of adult onset polycystic kidney disease re-evaluated. Kidney Int 26: 190-193. doi: 10.1038/ki.1984.154
|
| [5] |
Dell KM (2011) The spectrum of polycystic kidney disease in children. Adv Chronic Kidney Dis 18: 339-347. doi: 10.1053/j.ackd.2011.05.001
|
| [6] |
Romao EA, Moyses Neto M, Teixeira SR, et al. (2006) Renal and extrarenal manifestations of autosomal dominant polycystic kidney disease. Braz J Med Biol Res 39: 533-538. doi: 10.1590/S0100-879X2006000400014
|
| [7] |
Gabow PA (1993) Autosomal dominant polycystic kidney disease. N Engl J Med 329: 332-342. doi: 10.1056/NEJM199307293290508
|
| [8] | Huston J, 3rd, Torres VE, Sulivan PP, et al. (1993) Value of magnetic resonance angiography for the detection of intracranial aneurysms in autosomal dominant polycystic kidney disease. J Am Soc Nephrol 3: 1871-1877. |
| [9] | Abdul-Majeed S, Nauli SM (2011) Polycystic diseases in visceral organs. Obstet Gynecol Int 2011: 609370. |
| [10] | Ratnam S, Nauli SM (2010) Hypertension in Autosomal Dominant Polycystic Kidney Disease: A Clinical and Basic Science Perspective. Int J Nephrol Urol 2: 294-308. |
| [11] |
Zimmermann K (1898) Beiträge zur Kenntnis einiger Drüsen und Epithelien. Arch MikroskopAnat 52: 552-706. doi: 10.1007/BF02975837
|
| [12] | Nauli SM, Haymour HS, AbouAlaiwi WA, et al. (2011) Chapter 14: Primary Cilia are Mechanosensory Organelles in Vestibular Tissues. Mechanosensitivity and Mechanotransduction: ISBN: 978-990-481-9880-9881. |
| [13] | Jin X, Mohieldin AM, Muntean BS, et al. (2013) Cilioplasm is a cellular compartment for calcium signaling in response to mechanical and chemical stimuli. Cell Mol Life Sci. |
| [14] |
Garcia-Gonzalo FR, Reiter JF (2012) Scoring a backstage pass: mechanisms of ciliogenesis and ciliary access. J Cell Biol 197: 697-709. doi: 10.1083/jcb.201111146
|
| [15] |
Gherman A, Davis EE, Katsanis N (2006) The ciliary proteome database: an integrated community resource for the genetic and functional dissection of cilia. Nat Genet 38: 961-962. doi: 10.1038/ng0906-961
|
| [16] | Nauli SM, Jin X, Hierck BP (2011) The mechanosensory role of primary cilia in vascular hypertension. Int J Vasc Med 2011: 376281. |
| [17] |
Abdul-Majeed S, Nauli SM (2011) Calcium-mediated mechanisms of cystic expansion. Biochim Biophys Acta 1812: 1281-1290. doi: 10.1016/j.bbadis.2010.09.016
|
| [18] |
Nauli SM, Zhou J (2004) Polycystins and mechanosensation in renal and nodal cilia. Bioessays 26: 844-856. doi: 10.1002/bies.20069
|
| [19] |
Satir P, Pedersen LB, Christensen ST (2010) The primary cilium at a glance. J Cell Sci 123:499-503. doi: 10.1242/jcs.050377
|
| [20] |
Wood CR, Huang K, Diener DR, et al. (2013) The cilium secretes bioactive ectosomes. Curr Biol 23: 906-911. doi: 10.1016/j.cub.2013.04.019
|
| [21] |
Kaimori JY, Nagasawa Y, Menezes LF, et al. (2007) Polyductin undergoes notch-like processing and regulated release from primary cilia. Hum Mol Genet 16: 942-956. doi: 10.1093/hmg/ddm039
|
| [22] |
Qian F, Boletta A, Bhunia AK, et al. (2002) Cleavage of polycystin-1 requires the receptor for egg jelly domain and is disrupted by human autosomal-dominant polycystic kidney disease 1-associated mutations. Proc Natl Acad Sci U S A 99: 16981-16986. doi: 10.1073/pnas.252484899
|
| [23] |
Tanaka Y, Okada Y, Hirokawa N (2005) FGF-induced vesicular release of Sonic hedgehog and retinoic acid in leftward nodal flow is critical for left-right determination. Nature 435:172-177. doi: 10.1038/nature03494
|
| [24] |
Hogan MC, Manganelli L, Woollard JR, et al. (2009) Characterization of PKD proteinpositive exosome-like vesicles. J Am Soc Nephrol 20: 278-288. doi: 10.1681/ASN.2008060564
|
| [25] |
Praetorius HA, Spring KR (2003) Removal of the MDCK cell primary cilium abolishes flow sensing. J Membr Biol 191: 69-76. doi: 10.1007/s00232-002-1042-4
|
| [26] |
Nauli SM, Rossetti S, Kolb RJ, et al. (2006) Loss of polycystin-1 in human cyst-lining epithelia leads to ciliary dysfunction. J Am Soc Nephrol 17: 1015-1025. doi: 10.1681/ASN.2005080830
|
| [27] |
Nauli SM, Alenghat FJ, Luo Y, et al. (2003) Polycystins 1 and 2 mediate mechanosensation in the primary cilium of kidney cells. Nat Genet 33: 129-137. doi: 10.1038/ng1076
|
| [28] |
AbouAlaiwi WA, Takahashi M, Mell BR, et al. (2009) Ciliary polycystin-2 is a mechanosensitive calcium channel involved in nitric oxide signaling cascades. Circ Res 104:860-869. doi: 10.1161/CIRCRESAHA.108.192765
|
| [29] | Schwartz EA, Leonard ML, Bizios R, et al. (1997) Analysis and modeling of the primary cilium bending response to fluid shear. Am J Physiol 272: F132-138. |
| [30] | Downs ME, Nguyen AM, Herzog FA, et al. (2012) An experimental and computational analysis of primary cilia deflection under fluid flow. Comput Methods Biomech Biomed Engin. |
| [31] |
Resnick A, Hopfer U (2008) Mechanical stimulation of primary cilia. Front Biosci 13: 1665-1680. doi: 10.2741/2790
|
| [32] |
Rydholm S, Zwartz G, Kowalewski JM, et al. (2010) Mechanical properties of primary cilia regulate the response to fluid flow. Am J Physiol Renal Physiol 298: F1096-1102. doi: 10.1152/ajprenal.00657.2009
|
| [33] |
Forman JR, Qamar S, Paci E, et al. (2005) The remarkable mechanical strength of polycystin-1 supports a direct role in mechanotransduction. J Mol Biol 349: 861-871. doi: 10.1016/j.jmb.2005.04.008
|
| [34] |
Abdul-Majeed S, Nauli SM (2011) Dopamine receptor type 5 in the primary cilia has dual chemo- and mechano-sensory roles. Hypertension 58: 325-331. doi: 10.1161/HYPERTENSIONAHA.111.172080
|
| [35] | Hamon M, Doucet E, Lefevre K, et al. (1999) Antibodies and antisense oligonucleotide for probing the distribution and putative functions of central 5-HT6 receptors. Neuropsychopharmacology 21: 68S-76S. |
| [36] |
Handel M, Schulz S, Stanarius A, et al. (1999) Selective targeting of somatostatin receptor 3 to neuronal cilia. Neuroscience 89: 909-926. doi: 10.1016/S0306-4522(98)00354-6
|
| [37] |
Masyuk AI, Gradilone SA, Banales JM, et al. (2008) Cholangiocyte primary cilia are chemosensory organelles that detect biliary nucleotides via P2Y12 purinergic receptors. Am J Physiol Gastrointest Liver Physiol 295: G725-734. doi: 10.1152/ajpgi.90265.2008
|
| [38] |
Berbari NF, Johnson AD, Lewis JS, et al. (2008) Identification of ciliary localization sequences within the third intracellular loop of G protein-coupled receptors. Mol Biol Cell 19:1540-1547. doi: 10.1091/mbc.E07-09-0942
|
| [39] |
Rohatgi R, Milenkovic L, Scott MP (2007) Patched1 regulates hedgehog signaling at the primary cilium. Science 317: 372-376. doi: 10.1126/science.1139740
|
| [40] |
Corbit KC, Aanstad P, Singla V, et al. (2005) Vertebrate Smoothened functions at the primary cilium. Nature 437: 1018-1021. doi: 10.1038/nature04117
|
| [41] |
Kim E, Arnould T, Sellin LK, et al. (1999) The polycystic kidney disease 1 gene product modulates Wnt signaling. J Biol Chem 274: 4947-4953. doi: 10.1074/jbc.274.8.4947
|
| [42] |
Schneider L, Clement CA, Teilmann SC, et al. (2005) PDGFRalphaalpha signaling is regulated through the primary cilium in fibroblasts. Curr Biol 15: 1861-1866. doi: 10.1016/j.cub.2005.09.012
|
| [43] |
Soetedjo L, Glover DA, Jin H (2013) Targeting of vasoactive intestinal peptide receptor 2, VPAC2, a secretin family G-protein coupled receptor, to primary cilia. Biol Open 2: 686-694. doi: 10.1242/bio.20134747
|
| [44] |
Schneider L, Cammer M, Lehman J, et al. (2010) Directional cell migration and chemotaxis in wound healing response to PDGF-AA are coordinated by the primary cilium in fibroblasts. Cell Physiol Biochem 25: 279-292. doi: 10.1159/000276562
|
| [45] |
Jones TJ, Adapala RK, Geldenhuys WJ, et al. (2012) Primary cilia regulates the directional migration and barrier integrity of endothelial cells through the modulation of hsp27 dependent actin cytoskeletal organization. J Cell Physiol 227: 70-76. doi: 10.1002/jcp.22704
|
| [46] |
Jones TJ, Nauli SM (2012) Mechanosensory calcium signaling. Adv Exp Med Biol 740: 1001-1015. doi: 10.1007/978-94-007-2888-2_46
|
| [47] |
Choi YH, Suzuki A, Hajarnis S, et al. (2011) Polycystin-2 and phosphodiesterase 4C are components of a ciliary A-kinase anchoring protein complex that is disrupted in cystic kidney diseases. Proc Natl Acad Sci U S A 108: 10679-10684. doi: 10.1073/pnas.1016214108
|
| [48] |
Green JA, Gu C, Mykytyn K (2012) Heteromerization of ciliary G protein-coupled receptors in the mouse brain. PLoS One 7: e46304. doi: 10.1371/journal.pone.0046304
|
| [49] |
Marley A, Choy RW, von Zastrow M (2013) GPR88 reveals a discrete function of primary cilia as selective insulators of GPCR cross-talk. PLoS One 8: e70857. doi: 10.1371/journal.pone.0070857
|
| [50] | Kolb RJ, Nauli SM (2008) Ciliary dysfunction in polycystic kidney disease: an emerging model with polarizing potential. Front Biosci 13: 4451-4466. |
| [51] |
Nims N, Vassmer D, Maser RL (2003) Transmembrane domain analysis of polycystin-1, the product of the polycystic kidney disease-1 (PKD1) gene: evidence for 11 membranespanning domains. Biochemistry 42: 13035-13048. doi: 10.1021/bi035074c
|
| [52] |
Scheffers MS, van der Bent P, Prins F, et al. (2000) Polycystin-1, the product of the polycystic kidney disease 1 gene, co-localizes with desmosomes in MDCK cells. Hum Mol Genet 9: 2743-2750. doi: 10.1093/hmg/9.18.2743
|
| [53] |
Huan Y, van Adelsberg J (1999) Polycystin-1, the PKD1 gene product, is in a complex containing E-cadherin and the catenins. J Clin Invest 104: 1459-1468. doi: 10.1172/JCI5111
|
| [54] |
Yu S, Hackmann K, Gao J, et al. (2007) Essential role of cleavage of Polycystin-1 at G protein-coupled receptor proteolytic site for kidney tubular structure. Proc Natl Acad Sci U S A 104: 18688-18693. doi: 10.1073/pnas.0708217104
|
| [55] |
Low SH, Vasanth S, Larson CH, et al. (2006) Polycystin-1, STAT6, and P100 function in a pathway that transduces ciliary mechanosensation and is activated in polycystic kidney disease. Dev Cell 10: 57-69. doi: 10.1016/j.devcel.2005.12.005
|
| [56] |
Chauvet V, Tian X, Husson H, et al. (2004) Mechanical stimuli induce cleavage and nuclear translocation of the polycystin-1 C terminus. J Clin Invest 114: 1433-1443. doi: 10.1172/JCI21753
|
| [57] |
Lal M, Song X, Pluznick JL, et al. (2008) Polycystin-1 C-terminal tail associates with betacatenin and inhibits canonical Wnt signaling. Hum Mol Genet 17: 3105-3117. doi: 10.1093/hmg/ddn208
|
| [58] | Delmas P, Nauli SM, Li X, et al. (2004) Gating of the polycystin ion channel signaling complex in neurons and kidney cells. FASEB J 18: 740-742. |
| [59] |
Parnell SC, Magenheimer BS, Maser RL, et al. (1998) The polycystic kidney disease-1 protein, polycystin-1, binds and activates heterotrimeric G-proteins in vitro. Biochem Biophys Res Commun 251: 625-631. doi: 10.1006/bbrc.1998.9514
|
| [60] |
Parnell SC, Magenheimer BS, Maser RL, et al. (2002) Polycystin-1 activation of c-Jun Nterminal kinase and AP-1 is mediated by heterotrimeric G proteins. J Biol Chem 277: 19566-19572. doi: 10.1074/jbc.M201875200
|
| [61] |
Arnould T, Kim E, Tsiokas L, et al. (1998) The polycystic kidney disease 1 gene product mediates protein kinase C alpha-dependent and c-Jun N-terminal kinase-dependent activation of the transcription factor AP-1. J Biol Chem 273: 6013-6018. doi: 10.1074/jbc.273.11.6013
|
| [62] |
Le NH, van der Wal A, van der Bent P, et al. (2005) Increased activity of activator protein-1 transcription factor components ATF2, c-Jun, and c-Fos in human and mouse autosomal dominant polycystic kidney disease. J Am Soc Nephrol 16: 2724-2731. doi: 10.1681/ASN.2004110913
|
| [63] |
Shaulian E, Karin M (2002) AP-1 as a regulator of cell life and death. Nat Cell Biol 4: E131-136. doi: 10.1038/ncb0502-e131
|
| [64] |
Yu W, Kong T, Beaudry S, et al. (2010) Polycystin-1 protein level determines activity of the Galpha12/JNK apoptosis pathway. J Biol Chem 285: 10243-10251. doi: 10.1074/jbc.M109.070821
|
| [65] |
Puri S, Magenheimer BS, Maser RL, et al. (2004) Polycystin-1 activates the calcineurin/NFAT (nuclear factor of activated T-cells) signaling pathway. J Biol Chem 279:55455-55464. doi: 10.1074/jbc.M402905200
|
| [66] |
Distefano G, Boca M, Rowe I, et al. (2009) Polycystin-1 regulates extracellular signalregulated kinase-dependent phosphorylation of tuberin to control cell size through mTOR and its downstream effectors S6K and 4EBP1. Mol Cell Biol 29: 2359-2371. doi: 10.1128/MCB.01259-08
|
| [67] |
Shillingford JM, Murcia NS, Larson CH, et al. (2006) The mTOR pathway is regulated by polycystin-1, and its inhibition reverses renal cystogenesis in polycystic kidney disease. Proc Natl Acad Sci U S A 103: 5466-5471. doi: 10.1073/pnas.0509694103
|
| [68] |
Mochizuki T, Wu G, Hayashi T, et al. (1996) PKD2, a gene for polycystic kidney disease that encodes an integral membrane protein. Science 272: 1339-1342. doi: 10.1126/science.272.5266.1339
|
| [69] |
Cai Y, Maeda Y, Cedzich A, et al. (1999) Identification and characterization of polycystin-2, the PKD2 gene product. J Biol Chem 274: 28557-28565. doi: 10.1074/jbc.274.40.28557
|
| [70] |
Hanaoka K, Qian F, Boletta A, et al. (2000) Co-assembly of polycystin-1 and -2 produces unique cation-permeable currents. Nature 408: 990-994. doi: 10.1038/35050128
|
| [71] |
Qian F, Germino FJ, Cai Y, et al. (1997) PKD1 interacts with PKD2 through a probable coiled-coil domain. Nat Genet 16: 179-183. doi: 10.1038/ng0697-179
|
| [72] |
Nauli SM, Jin X, AbouAlaiwi WA, et al. (2013) Non-motile primary cilia as fluid shear stress mechanosensors. Methods Enzymol 525: 1-20. doi: 10.1016/B978-0-12-397944-5.00001-8
|
| [73] |
Liu W, Murcia NS, Duan Y, et al. (2005) Mechanoregulation of intracellular Ca2+ concentration is attenuated in collecting duct of monocilium-impaired orpk mice. Am J Physiol Renal Physiol 289: F978-988. doi: 10.1152/ajprenal.00260.2004
|
| [74] |
Praetorius HA, Spring KR (2001) Bending the MDCK cell primary cilium increases intracellular calcium. J Membr Biol 184: 71-79. doi: 10.1007/s00232-001-0075-4
|
| [75] |
Xu C, Shmukler BE, Nishimura K, et al. (2009) Attenuated, flow-induced ATP release contributes to absence of flow-sensitive, purinergic Cai2+ signaling in human ADPKD cyst epithelial cells. Am J Physiol Renal Physiol 296: F1464-1476. doi: 10.1152/ajprenal.90542.2008
|
| [76] |
Li Y, Wright JM, Qian F, et al. (2005) Polycystin 2 interacts with type I inositol 1,4,5- trisphosphate receptor to modulate intracellular Ca2+ signaling. J Biol Chem 280: 41298-41306. doi: 10.1074/jbc.M510082200
|
| [77] |
Li Y, Santoso NG, Yu S, et al. (2009) Polycystin-1 interacts with inositol 1,4,5-trisphosphate receptor to modulate intracellular Ca2+ signaling with implications for polycystic kidney disease. J Biol Chem 284: 36431-36441. doi: 10.1074/jbc.M109.068916
|
| [78] |
Praetorius HA, Leipziger J (2009) Released nucleotides amplify the cilium-dependent, flowinduced [Ca2+]i response in MDCK cells. Acta Physiol (Oxf) 197: 241-251. doi: 10.1111/j.1748-1716.2009.02002.x
|
| [79] |
Praetorius HA, Praetorius J, Nielsen S, et al. (2004) Beta1-integrins in the primary cilium of MDCK cells potentiate fibronectin-induced Ca2+ signaling. Am J Physiol Renal Physiol 287: F969-978. doi: 10.1152/ajprenal.00096.2004
|
| [80] |
Eggenschwiler JT, Anderson KV (2007) Cilia and developmental signaling. Annu Rev Cell Dev Biol 23: 345-373. doi: 10.1146/annurev.cellbio.23.090506.123249
|
| [81] |
Lum L, Beachy PA (2004) The Hedgehog response network: sensors, switches, and routers. Science 304: 1755-1759. doi: 10.1126/science.1098020
|
| [82] | Mohieldin AM, Upadhyay VS, Ong ACM, et al. (2013) Autosomal Dominant Polycystic Kidney Disease: Pathophysiology and Treatment. Autosomal Dominant Disorders: New Research ISBN: 978-1-62808-761-1: 6x9 - (NBC-R). |
| [83] |
Nauli SM (2011) An ACE inhibitor improves vascular outcomes in a PKD model. Am J Physiol Renal Physiol 301: F958. doi: 10.1152/ajprenal.00489.2011
|
| [84] |
Andersson M, Karlsson L, Svensson PA, et al. (2005) Differential global gene expression response patterns of human endothelium exposed to shear stress and intraluminal pressure. J Vasc Res 42: 441-452. doi: 10.1159/000087983
|
| [85] |
Masuda H, Zhuang YJ, Singh TM, et al. (1999) Adaptive remodeling of internal elastic lamina and endothelial lining during flow-induced arterial enlargement. Arterioscler Thromb Vasc Biol 19: 2298-2307. doi: 10.1161/01.ATV.19.10.2298
|
| [86] |
Pries AR, Secomb TW, Gaehtgens P (1995) Design principles of vascular beds. Circ Res 77:1017-1023. doi: 10.1161/01.RES.77.5.1017
|
| [87] |
Ohura N, Yamamoto K, Ichioka S, et al. (2003) Global analysis of shear stress-responsive genes in vascular endothelial cells. J Atheroscler Thromb 10: 304-313. doi: 10.5551/jat.10.304
|
| [88] | Langille BL, Bendeck MP, Keeley FW (1989) Adaptations of carotid arteries of young and mature rabbits to reduced carotid blood flow. Am J Physiol 256: H931-939. |
| [89] |
Langille BL, O'Donnell F (1986) Reductions in arterial diameter produced by chronic decreases in blood flow are endothelium-dependent. Science 231: 405-407. doi: 10.1126/science.3941904
|
| [90] |
Huang C, Holfeld J, Schaden W, et al. (2013) Mechanotherapy: revisiting physical therapy and recruiting mechanobiology for a new era in medicine. Trends Mol Med 19: 555-564. doi: 10.1016/j.molmed.2013.05.005
|
| [91] |
Nauli SM, Kawanabe Y, Kaminski JJ, et al. (2008) Endothelial cilia are fluid shear sensors that regulate calcium signaling and nitric oxide production through polycystin-1. Circulation 117: 1161-1171. doi: 10.1161/CIRCULATIONAHA.107.710111
|
| [92] |
Hierck BP, Van der Heiden K, Alkemade FE, et al. (2008) Primary cilia sensitize endothelial cells for fluid shear stress. Dev Dyn 237: 725-735. doi: 10.1002/dvdy.21472
|
| [93] |
Wang N, Miao H, Li YS, et al. (2006) Shear stress regulation of Kruppel-like factor 2 expression is flow pattern-specific. Biochem Biophys Res Commun 341: 1244-1251. doi: 10.1016/j.bbrc.2006.01.089
|
| [94] | Boon RA, Horrevoets AJ (2009) Key transcriptional regulators of the vasoprotective effects of shear stress. Hamostaseologie 29: 39-40, 41-33. |
| [95] |
Dekker RJ, van Thienen JV, Rohlena J, et al. (2005) Endothelial KLF2 links local arterial shear stress levels to the expression of vascular tone-regulating genes. Am J Pathol 167:609-618. doi: 10.1016/S0002-9440(10)63002-7
|
| [96] |
SenBanerjee S, Lin Z, Atkins GB, et al. (2004) KLF2 Is a novel transcriptional regulator of endothelial proinflammatory activation. J Exp Med 199: 1305-1315. doi: 10.1084/jem.20031132
|
| [97] |
Abdul-Majeed S, Moloney BC, Nauli SM (2012) Mechanisms regulating cilia growth and cilia function in endothelial cells. Cell Mol Life Sci 69: 165-173. doi: 10.1007/s00018-011-0744-0
|
| [98] |
Clement DL, Mally S, Stock C, et al. (2013) PDGFRalpha signaling in the primary cilium regulates NHE1-dependent fibroblast migration via coordinated differential activity of MEK1/2-ERK1/2-p90RSK and AKT signaling pathways. J Cell Sci 126: 953-965. doi: 10.1242/jcs.116426
|
| [99] |
AbouAlaiwi WA, Ratnam S, Booth RL, et al. (2011) Endothelial cells from humans and mice with polycystic kidney disease are characterized by polyploidy and chromosome segregation defects through survivin down-regulation. Hum Mol Genet 20: 354-367. doi: 10.1093/hmg/ddq470
|
| [100] | AbouAlaiwi WA, Rodriguez I, Nauli SM (2012) Spectral karyotyping to study chromosome abnormalities in humans and mice with polycystic kidney disease. J Vis Exp. |
| [101] | Aboualaiwi WA, Muntean BS, Ratnam S, et al. (2013) Survivin-Induced Abnormal Ploidy Contributes to Cystic Kidney and Aneurysm Formation. Circulation. |
| [102] |
Zaragoza C, Marquez S, Saura M (2012) Endothelial mechanosensors of shear stress as regulators of atherogenesis. Curr Opin Lipidol 23: 446-452. doi: 10.1097/MOL.0b013e328357e837
|
| [103] |
Van der Heiden K, Hierck BP, Krams R, et al. (2008) Endothelial primary cilia in areas of disturbed flow are at the base of atherosclerosis. Atherosclerosis 196: 542-550. doi: 10.1016/j.atherosclerosis.2007.05.030
|
| [104] |
Bystrevskaya VB, Lichkun VV, Antonov AS, et al. (1988) An ultrastructural study of centriolar complexes in adult and embryonic human aortic endothelial cells. Tissue Cell 20:493-503. doi: 10.1016/0040-8166(88)90052-3
|
| [105] |
Iomini C, Tejada K, Mo W, et al. (2004) Primary cilia of human endothelial cells disassemble under laminar shear stress. J Cell Biol 164: 811-817. doi: 10.1083/jcb.200312133
|
| [106] |
Abou Alaiwi WA, Lo ST, Nauli SM (2009) Primary cilia: highly sophisticated biological sensors. Sensors (Basel) 9: 7003-7020. doi: 10.3390/s90907003
|
| [107] |
Teilmann SC, Christensen ST (2005) Localization of the angiopoietin receptors Tie-1 and Tie-2 on the primary cilia in the female reproductive organs. Cell Biol Int 29: 340-346. doi: 10.1016/j.cellbi.2005.03.006
|
| [108] |
Hazzard TM, Molskness TA, Chaffin CL, et al. (1999) Vascular endothelial growth factor (VEGF) and angiopoietin regulation by gonadotrophin and steroids in macaque granulosa cells during the peri-ovulatory interval. Mol Hum Reprod 5: 1115-1121. doi: 10.1093/molehr/5.12.1115
|
| [109] |
Kim I, Kim HG, So JN, et al. (2000) Angiopoietin-1 regulates endothelial cell survival through the phosphatidylinositol 3'-Kinase/Akt signal transduction pathway. Circ Res 86: 24-29. doi: 10.1161/01.RES.86.1.24
|
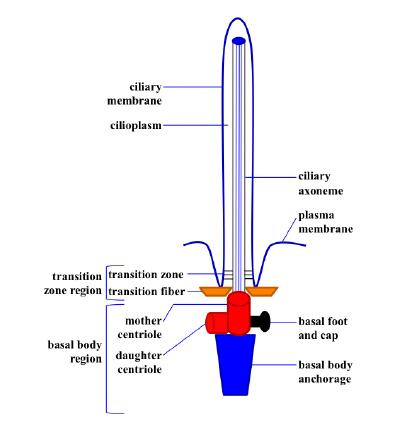
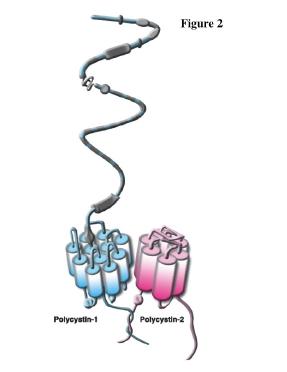
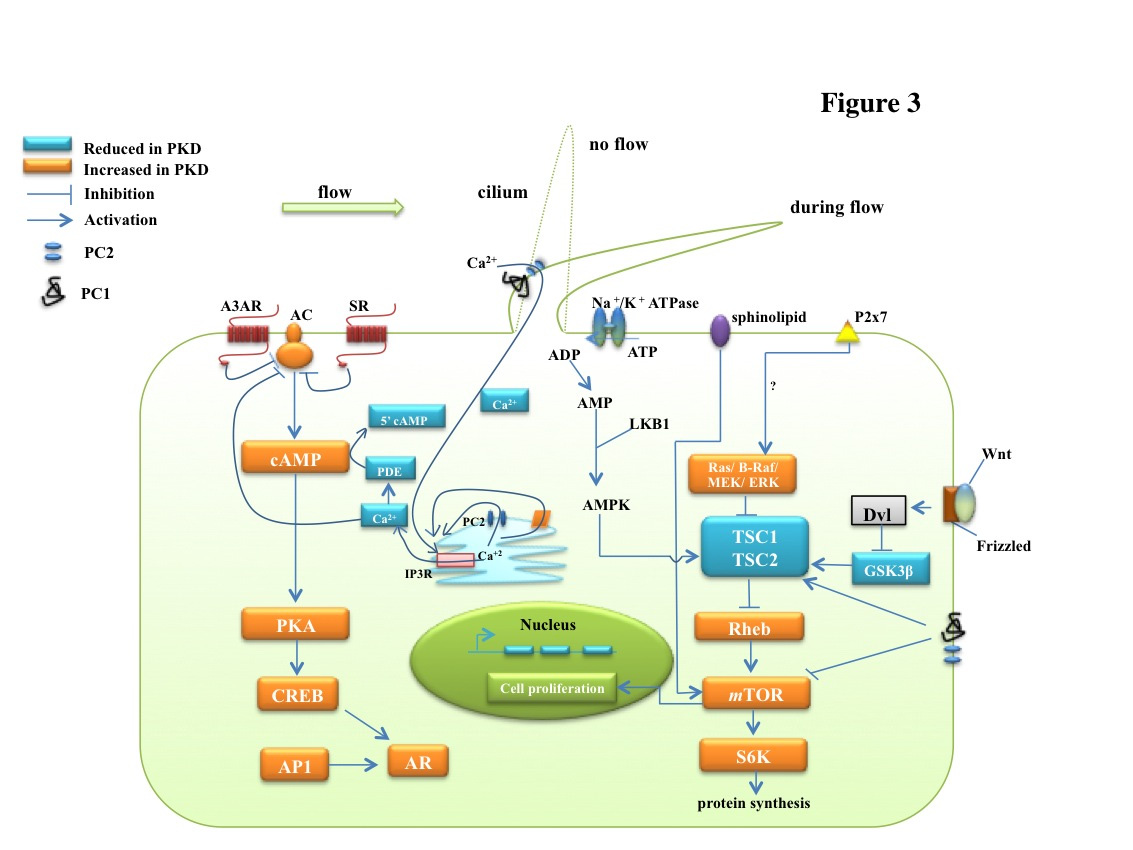
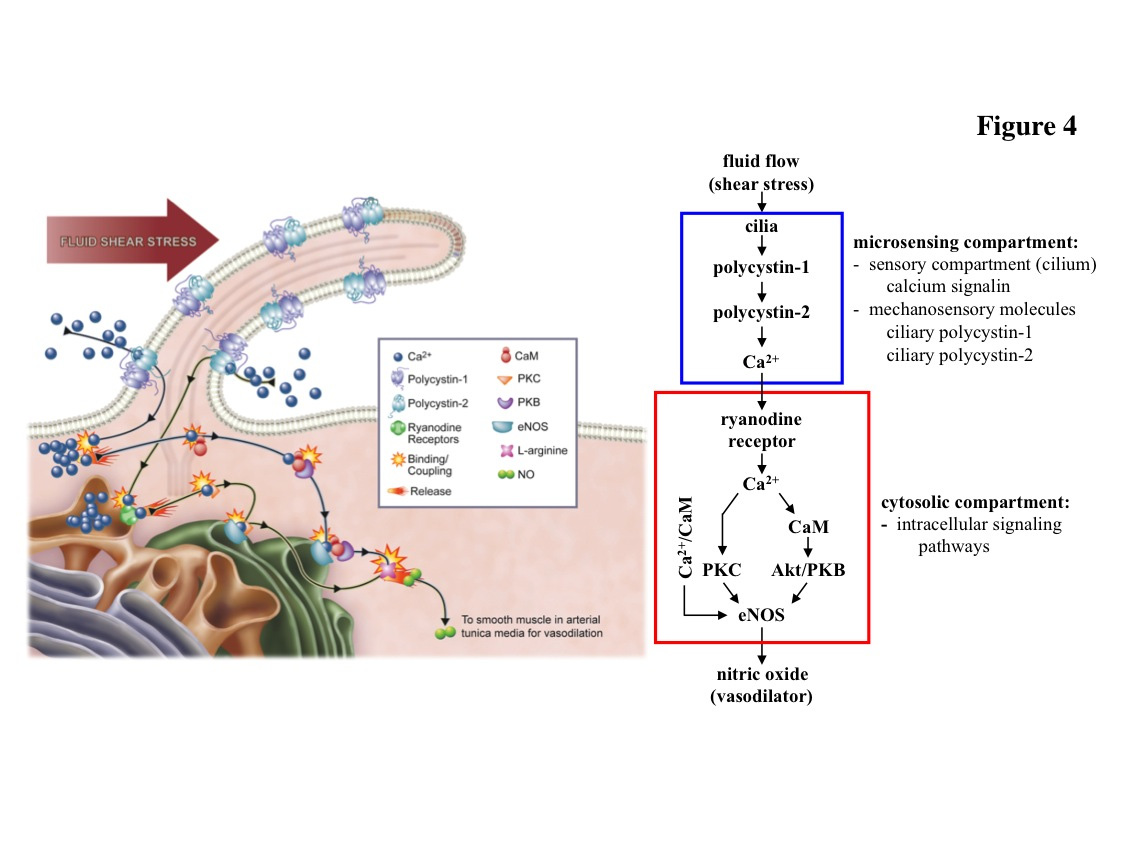
Figures(4)

Sarmed H. Kathem, Ashraf M. Mohieldin, Surya M. Nauli. The Roles of Primary cilia in Polycystic Kidney Disease[J]. AIMS Molecular Science, 2014, 1(1): 27-46. doi: 10.3934/molsci.2013.1.27







 DownLoad:
DownLoad: