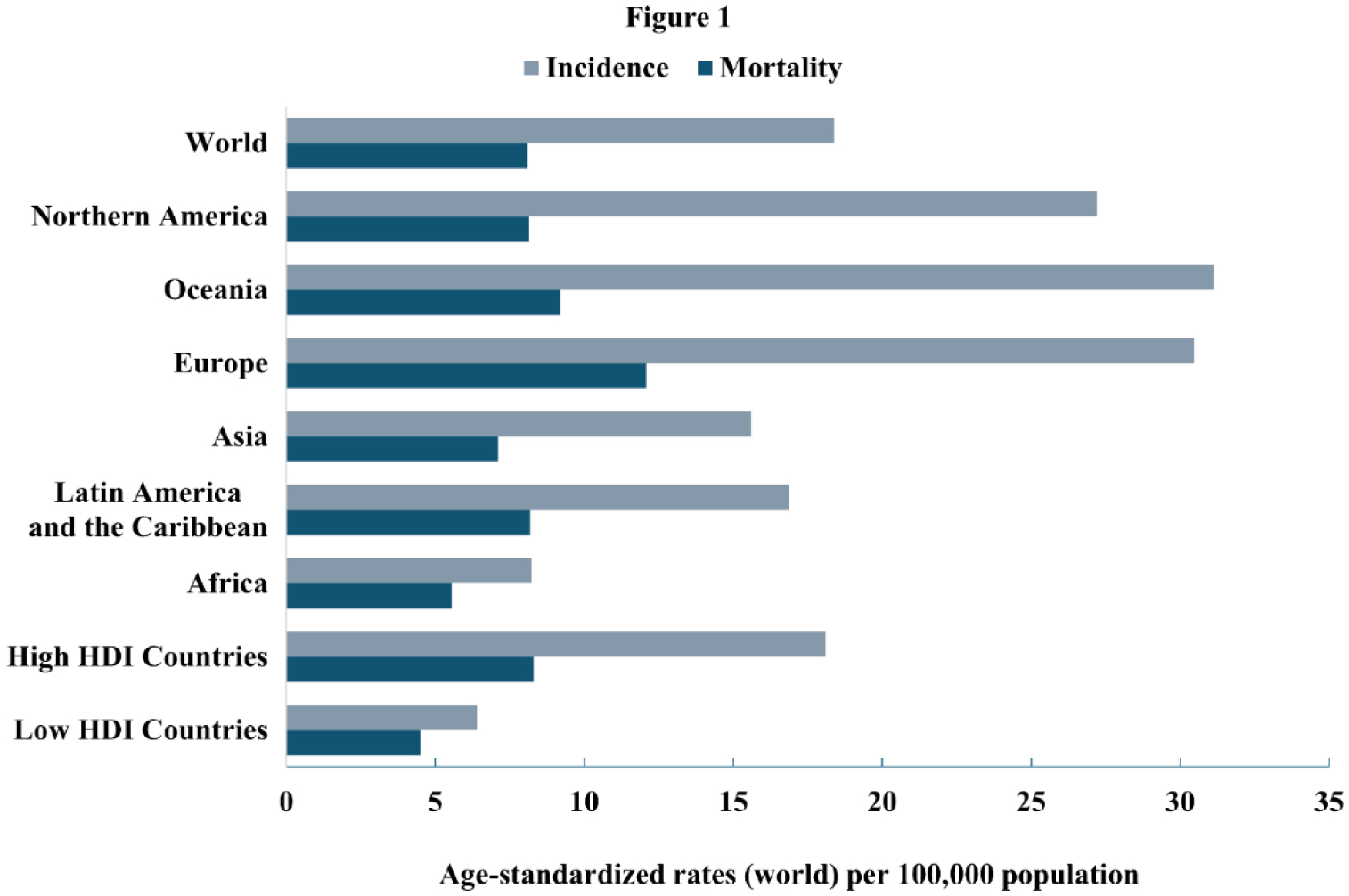
Colorectal cancer (CRC), a cancer of the large intestine and rectum, has one of the highest incidence and mortality rates, with incidences expected to nearly double in the next 25 years. The development of CRC may be caused by a collection of mutations via the chromosomal instability pathway, which results in approximately 70% of sporadic CRC cases, or the microsatellite instability pathway. At the same time, there are various modifiable and non-modifiable risk factors that increase one's chances of developing a CRC causing mutation via these pathways. There are three widely used treatments for CRC: surgical resection, chemotherapy, and immunotherapy. Surgical resection remains the standard for CRC that has not metastasized with various options available such as open surgery, laparoscopy, and robotic surgery. In recent years, there has also been a plethora of new research into chemotherapies and immunotherapies as important treatments for metastatic or complicated CRC cases. In this review, we highlight the pathogenesis, risk factors, and treatments for CRC, while also providing a short summary of the diagnosis. Such a study will be helpful for clinicians to better manage patients with CRC.
Citation: Derek A. Corica, Dylan M. Schaap, Trenton G. Mayberry, Braydon C. Cowan, Mark R. Wakefield, Yujiang Fang. New depths: exploring the current landscape of colorectal cancer[J]. AIMS Medical Science, 2025, 12(1): 105-123. doi: 10.3934/medsci.2025008
Colorectal cancer (CRC), a cancer of the large intestine and rectum, has one of the highest incidence and mortality rates, with incidences expected to nearly double in the next 25 years. The development of CRC may be caused by a collection of mutations via the chromosomal instability pathway, which results in approximately 70% of sporadic CRC cases, or the microsatellite instability pathway. At the same time, there are various modifiable and non-modifiable risk factors that increase one's chances of developing a CRC causing mutation via these pathways. There are three widely used treatments for CRC: surgical resection, chemotherapy, and immunotherapy. Surgical resection remains the standard for CRC that has not metastasized with various options available such as open surgery, laparoscopy, and robotic surgery. In recent years, there has also been a plethora of new research into chemotherapies and immunotherapies as important treatments for metastatic or complicated CRC cases. In this review, we highlight the pathogenesis, risk factors, and treatments for CRC, while also providing a short summary of the diagnosis. Such a study will be helpful for clinicians to better manage patients with CRC.
Colorectal cancer
Chromosomal instability pathway
Microsatellite instability pathway
Laparoscopic-assisted surgery
Robot assisted surgery
Immunotherapy
Immune-related adverse events

| [1] | Kochanek KD, Murphy SL, Xu JQ, et al. (2024) Mortality in the United States, 2022. NCHS Data Brief, no 492 . Hyattsville, MD: National Center for Health Statistics. 2024. https://dx.doi.org/10.15620/cdc:135850. |
| [2] | Global Cancer Observatory, World Health Organization, Colon and Rectum. Accessed 12 March 2024. Available from: https://gco.iarc.fr/today/en/fact-sheets-cancers |
| [3] |
Bray F, Laversanne M, Sung H, et al. (2024) Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 74: 229-263. https://doi.org/10.3322/caac.21834 
|
| [4] | Ferlay J, Ervik M, Lam F, et al. (2024) Global Cancer Observatory: Cancer Today (version 1.1). Lyon, France: International Agency for Research on Cancer. Available from: https://gco.iarc.who.int/today. |
| [5] |
Morgan E, Arnold M, Gini A, et al. (2023) Global burden of colorectal cancer in 2020 and 2040: incidence and mortality estimates from GLOBOCAN. Gut 72: 338-344. https://doi.org/10.1136/gutjnl-2022-327736
|
| [6] |
Sullivan BA, Noujaim M, Roper J (2022) Cause, epidemiology, and histology of polyps and pathways to colorectal cancer. Gastrointest Endosc Clin N Am 32: 177-194. https://doi.org/10.1016/j.giec.2021.12.001
|
| [7] |
Nguyen LH, Goel A, Chung DC (2020) Pathways of colorectal carcinogenesis. Gastroenterology 158: 291-302. https://doi.org/10.1053/j.gastro.2019.08.059
|
| [8] |
Simon K (2016) Colorectal cancer development and advances in screening. Clin Interv Aging 11: 967-976. https://doi.org/10.2147/CIA.S109285
|
| [9] |
Pickhardt PJ, Kim DH (2009) Colorectal cancer screening with CT colonography: key concepts regarding polyp prevalence, size, histology, morphology, and natural history. AJR Am J Roentgenol 193: 40-46. https://doi.org/10.2214/AJR.08.1709
|
| [10] |
Pino MS, Chung DC (2010) The chromosomal instability pathway in colon cancer. Gastroenterology 138: 2059-2072. https://doi.org/10.1053/j.gastro.2009.12.065
|
| [11] |
Powell SM, Zilz N, Beazer-Barclay Y, et al. (1992) APC mutations occur early during colorectal tumorigenesis. Nature 359: 235-237. https://doi.org/10.1038/359235a0
|
| [12] |
Rim EY, Clevers H, Nusse R (2022) The Wnt pathway: from signaling mechanisms to synthetic modulators. Annu Rev Biochem 91: 571-598. https://doi.org/10.1146/annurev-biochem-040320-103615
|
| [13] |
Dienstmann R, Connor K, Byrne AT (2020) COLOSSUS consortium. Precision therapy in RAS mutant colorectal cancer. Gastroenterology 158: 806-811. https://doi.org/10.1053/j.gastro.2019.12.051
|
| [14] |
Fernández-Medarde A, Santos E (2011) Ras in cancer and developmental diseases. Genes Cancer 2: 344-358. https://doi.org/10.1177/1947601911411084
|
| [15] |
Simanshu DK, Nissley DV, McCormick F (2017) RAS proteins and their regulators in human disease. Cell 170: 17-33. https://doi.org/10.1016/j.cell.2017.06.009
|
| [16] |
Zhu G, Pei L, Xia H, et al. (2021) Role of oncogenic KRAS in the prognosis, diagnosis and treatment of colorectal cancer. Mol Cancer 20: 143. https://doi.org/10.1186/s12943-021-01441-4
|
| [17] |
McLellan EA, Owen RA, Stepniewska KA, et al. (1993) Lemoine NR. High frequency of K-ras mutations in sporadic colorectal adenomas. Gut 34: 392-396. https://doi.org/10.1136/gut.34.3.392
|
| [18] |
Naccarati A, Polakova V, Pardini B, et al. (2012) Mutations and polymorphisms in TP53 gene--an overview on the role in colorectal cancer. Mutagenesis 27: 211-218. https://doi.org/10.1093/mutage/ger067
|
| [19] |
Fischer M (2017) Census and evaluation of p53 target genes. Oncogene 36: 3943-3956. https://doi.org/10.1038/onc.2016.502
|
| [20] |
Lin D, Shields MT, Ullrich SJ, et al. (1992) Growth arrest induced by wild-type p53 protein blocks cells prior to or near the restriction point in late G1 phase. Proc Natl Acad Sci USA 89: 9210-9214. https://doi.org/10.1073/pnas.89.19.9210
|
| [21] |
Engeland K (2022) Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ 29: 946-960. https://doi.org/10.1038/s41418-022-00988-z
|
| [22] |
Iacopetta B, Russo A, Bazan V, et al. (2006) Functional categories of TP53 mutation in colorectal cancer: results of an International Collaborative Study. Ann Oncol 217: 842-847. https://doi.org/10.1093/annonc/mdl035
|
| [23] |
Leslie A, Carey FA, Pratt NR (2002) The colorectal adenoma-carcinoma sequence. Br J Surg 89: 845-860. https://doi.org/10.1046/j.1365-2168.2002.02120.x
|
| [24] |
Westra JL, Schaapveld M, Hollema H, et al. (2005) Determination of TP53 mutation is more relevant than microsatellite instability status for the prediction of disease-free survival in adjuvant-treated stage III colon cancer patients. J Clin Oncol 23: 5635-5643. https://doi.org/10.1200/JCO.2005.04.096
|
| [25] |
Söreide K, Janssen EA, Söiland H, et al. (2006) Microsatellite instability in colorectal cancer. Br J Surg 93: 395-406.
|
| [26] | De' Angelis GL, Bottarelli L, Azzoni C, et al. (2018) Microsatellite instability in colorectal cancer. Acta Biomed 89: 97-101. https://doi.org/10.23750/abm.v89i9-S.7960 |
| [27] | Parsons R, Li GM, Longley MJ, et al. (1993) Hypermutability and mismatch repair deficiency in RER+ tumor cells. Cell 75: 1227-1236. https://doi.org/10.1016/0092-8674(93)90331-j |
| [28] |
Hendriks YM, de Jong AE, Morreau H, et al. (2006) Diagnostic approach and management of Lynch syndrome (hereditary nonpolyposis colorectal carcinoma): a guide for clinicians. CA Cancer J Clin 56: 213-225. https://doi.org/10.3322/canjclin.56.4.213
|
| [29] |
Iacopetta B, Grieu F, Amanuel B (2010) Microsatellite instability in colorectal cancer. Asia Pac J Clin Oncol 6: 260-269. https://doi.org/10.1111/j.1743-7563.2010.01335.x
|
| [30] |
Sninsky JA, Shore BM, Lupu GV (2022) Crockett SD. Risk factors for colorectal polyps and cancer. Gastrointest Endosc Clin N Am 32: 195-213. https://doi.org/10.1016/j.giec.2021.12.008
|
| [31] |
Lewandowska A, Rudzki G, Lewandowski T (2022) Risk factors for the diagnosis of colorectal cancer. Cancer Control 29. https://doi.org/10.1177/10732748211056692
|
| [32] |
Betesh AL, Schnoll-Sussman FH (2021) Colorectal cancer screening in the elderly. Clin Geriatr Med 37: 173-183. https://doi.org/10.1016/j.cger.2020.08.012
|
| [33] | Gupta S (2022) Screening for colorectal cancer. Hematol Oncol Clin North Am 36: 393-414. https://doi.org/10.1016/j.hoc.2022.02.001 |
| [34] |
Mehraban Far P, Alshahrani A, Yaghoobi M (2019) Quantitative risk of positive family history in developing colorectal cancer: A meta-analysis. World J Gastroenterol 25: 4278-4291. https://doi.org/10.3748/wjg.v25.i30.4278
|
| [35] | Mármol I, Sánchez-de-Diego C, Pradilla Dieste A, et al. (2017) Colorectal carcinoma: A general overview and future perspectives in colorectal cancer. Int J Mol Sci 18: 197. https://doi.org/10.3390/ijms18010197 |
| [36] |
Siegel RL, Torre LA, Soerjomataram I, et al. (2019) Global patterns and trends in colorectal cancer incidence in young adults. Gut 68: 2179-2185. https://doi.org/10.1136/gutjnl-2019-319511
|
| [37] |
Bailey CE, Hu CY, You YN, et al. (2015) Increasing disparities in the age-related incidences of colon and rectal cancers in the United States, 1975–2010. JAMA Surg 150: 17-22. https://doi.org/10.1001/jamasurg.2014.1756
|
| [38] |
O'Sullivan DE, Sutherland RL, Town S, et al. (2022) Risk factors for early-onset colorectal cancer: A systematic review and Meta-analysis. Clin Gastroenterol Hepatol 20: 1229-1240.e5. https://doi.org/10.1016/j.cgh.2021.01.037
|
| [39] |
Ionescu VA, Gheorghe G, Bacalbasa N, et al. (2023) Colorectal cancer: from risk factors to oncogenesis. Medicina 59: 1646. https://doi.org/10.3390/medicina59091646
|
| [40] |
Zhou X, Wang L, Xiao J, et al. (2022) Alcohol consumption, DNA methylation and colorectal cancer risk: Results from pooled cohort studies and Mendelian randomization analysis. Int J Cancer 151: 83-94. https://doi.org/10.1002/ijc.33945
|
| [41] |
Farvid MS, Sidahmed E, Spence ND, et al. (2021) Consumption of red meat and processed meat and cancer incidence: a systematic review and meta-analysis of prospective studies. Eur J Epidemiol 36: 937-951. https://doi.org/10.1007/s10654-021-00741-9
|
| [42] |
Ladabaum U, Dominitz JA, Kahi C, et al. (2020) Strategies for colorectal cancer screening. Gastroenterology 158: 418-432. https://doi.org/10.1053/j.gastro.2019.06.043
|
| [43] |
Wu Z, Li Y, Zhang Y, et al. (2020) Colorectal cancer screening methods and molecular markers for early detection. Technol Cancer Res Treat 19: 1533033820980426. https://doi.org/10.1177/1533033820980426
|
| [44] |
Schreuders EH, Ruco A, Rabeneck L, et al. (2015) Colorectal cancer screening: a global overview of existing programmes. Gut 64: 1637-1649. https://doi.org/10.1136/gutjnl-2014-309086
|
| [45] |
Shaukat A, Levin TR (2022) Current and future colorectal cancer screening strategies. Nat Rev Gastroenterol Hepatol 19: 521-531. https://doi.org/10.1038/s41575-022-00612-y
|
| [46] |
Potter NT, Hurban P, White MN, et al. (2014) Validation of a real-time PCR-based qualitative assay for the detection of methylated SEPT9 DNA in human plasma. Clin Chem 60: 1183-1191. https://doi.org/10.1373/clinchem.2013.221044
|
| [47] |
Cohen JD, Li L, Wang Y, et al. (2018) Detection and localization of surgically resectable cancers with a multi-analyte blood test. Science 359: 926-930. https://doi.org/10.1126/science.aar3247
|
| [48] |
Barnell EK, Wurtzler EM, La Rocca J, et al. (2023) Multitarget stool RNA test for colorectal cancer screening. JAMA 330: 1760-1768. https://doi.org/10.1001/jama.2023.22231
|
| [49] |
Mahmoud NN (2022) Colorectal cancer: preoperative evaluation and staging. Surg Oncol Clin N Am 31: 127-141. https://doi.org/10.1016/j.soc.2021.12.001
|
| [50] |
Nagtegaal ID, Quirke P, Schmoll HJ (2011) Has the new TNM classification for colorectal cancer improved care?. Nat Rev Clin Oncol 9: 119-123. https://doi.org/10.1038/nrclinonc.2011.157
|
| [51] |
Chen K, Collins G, Wang H, et al. (2021) Pathological features and prognostication in colorectal cancer. Curr Oncol 28: 5356-5383. https://doi.org/10.3390/curroncol28060447
|
| [52] |
Carlsen L, Huntington KE, El-Deiry WS (2022) Immunotherapy for Colorectal Cancer: Mechanisms and Predictive Biomarkers. Cancers 14: 1028. https://doi.org/10.3390/cancers14041028
|
| [53] |
Zhao W, Jin L, Chen P, et al. (2022) Colorectal cancer immunotherapy-Recent progress and future directions. Cancer Lett 545: 215816. https://doi.org/10.1016/j.canlet.2022.215816
|
| [54] |
Shinji S, Yamada T, Matsuda A, et al. (2022) Recent Advances in the Treatment of Colorectal Cancer: A Review. J Nippon Med Sch 89: 246-254.
|
| [55] |
Merchant J, McArthur D, Ferguson H, et al. (2021) Concepts and prospects of minimally invasive colorectal cancer surgery. Clin Radiol 76: 889-895. https://doi.org/10.1016/j.crad.2021.09.013
|
| [56] | Vilsan J, Maddineni SA, Ahsan N, et al. (2023) Open, Laparoscopic, and Robotic Approaches to Treat Colorectal Cancer: A Comprehensive Review of Literature. Cureus 15: e38956. https://doi.org/10.7759/cureus.38956 |
| [57] |
Zhang M, Liu Z, Wang X (2022) Is natural orifice specimen extraction surgery the future direction of minimally invasive colorectal surgery?. Surg Open Sci 10: 106-110. https://doi.org/10.1016/j.sopen.2022.08.001
|
| [58] |
Song XJ, Liu ZL, Zeng R, et al. (2019) A meta-analysis of laparoscopic surgery versus conventional open surgery in the treatment of colorectal cancer. Medicine 98: e15347. https://doi.org/10.1097/MD.0000000000015347
|
| [59] |
Luo W, Wu M, Chen Y (2022) Laparoscopic versus open surgery for elderly patients with colorectal cancer: a systematic review and meta-analysis of matched studies. ANZ J Surg 92: 2003-2017. https://doi.org/10.1111/ans.17972
|
| [60] |
Yang L, Fang C, Bi T, et al. (2023) Efficacy of robot-assisted vs. laparoscopy surgery in the treatment of colorectal cancer: A systematic review and meta-analysis. Clin Res Hepatol Gastroenterol 47: 102176. https://doi.org/10.1016/j.clinre.2023.102176
|
| [61] |
Huang Z, Huang S, Huang Y, et al. (2023) Comparison of robotic-assisted versus conventional laparoscopic surgery in colorectal cancer resection: a systemic review and meta-analysis of randomized controlled trials. Front Oncol 13: 1273378. https://doi.org/10.3389/fonc.2023.1273378
|
| [62] |
Zhao Z, Gu J (2022) Open surgery in the era of minimally invasive surgery. Chin J Cancer Res 34: 63-65. https://doi.org/10.21147/j.issn.1000-9604.2022.01.06
|
| [63] |
Biller LH, Schrag D (2021) Diagnosis and treatment of metastatic colorectal cancer: A review. JAMA 325: 669-685. https://doi.org/10.1001/jama.2021.0106
|
| [64] | European Medicines AgencyHomepage. Accessed March 8, 2025. Available from: https://www.ema.europa.eu/en/homepage |
| [65] | National Cancer InstituteColorectal cancer treatment drugs. Accessed March 8, 2025. Available from: https://www.cancer.gov/about-cancer/treatment/drugs/colorectal |
| [66] |
Kuipers EJ, Grady WM, Lieberman D, et al. (2015) Colorectal cancer. Nat Rev Dis Primers 1: 15065. https://doi.org/10.1038/nrdp.2015.65
|
| [67] |
Glimelius B, Stintzing S, Marshall J, et al. (2021) Metastatic colorectal cancer: Advances in the folate-fluoropyrimidine chemotherapy backbone. Cancer Treat Rev 98: 102218. https://doi.org/10.1016/j.ctrv.2021.102218
|
| [68] |
Shin AE, Giancotti FG, Rustgi AK (2023) Metastatic colorectal cancer: mechanisms and emerging therapeutics. Trends Pharmacol Sci 44: 222-236. https://doi.org/10.1016/j.tips.2023.01.003
|
| [69] |
Colucci G, Gebbia V, Paoletti G, et al. (2005) Phase III randomized trial of FOLFIRI versus FOLFOX4 in the treatment of advanced colorectal cancer: a multicenter study of the Gruppo Oncologico Dell'Italia Meridionale. J Clin Oncol 23: 4866-4875. https://doi.org/10.1200/JCO.2005.07.113
|
| [70] |
Nelson MA, Shetty S, Kulakodlu M, et al. (2011) A comparison of mortality and costs associated with FOLFOX versus FOLFIRI in stage IV colorectal cancer. J Med Econ 14: 179-186. https://doi.org/10.3111/13696998.2011.556693
|
| [71] |
Van Cutsem E, Köhne CH, Hitre E, et al. (2009) Cetuximab and chemotherapy as initial treatment for metastatic colorectal cancer. N Engl J Med 360: 1408-1417. https://doi.org/10.1056/NEJMoa0805019
|
| [72] |
Douillard JY, Siena S, Cassidy J, et al. (2010) Randomized, phase III trial of panitumumab with infusional fluorouracil, leucovorin, and oxaliplatin (FOLFOX4) versus FOLFOX4 alone as first-line treatment in patients with previously untreated metastatic colorectal cancer: the PRIME study. J Clin Oncol 28: 4697-4705. https://doi.org/10.1200/JCO.2009.27.4860
|
| [73] |
Itatani Y, Kawada K, Yamamoto T, et al. (2018) Resistance to anti-angiogenic therapy in Cancer-Alterations to Anti-VEGF Pathway. Int J Mol Sci 19: 1232. https://doi.org/10.3390/ijms19041232
|
| [74] |
Diaz LA, Shiu KK, Kim TW, et al. (2022) Pembrolizumab versus chemotherapy for microsatellite instability-high or mismatch repair-deficient metastatic colorectal cancer (KEYNOTE-177): final analysis of a randomised, open-label, phase 3 study. Lancet Oncol 23: 659-670. https://doi.org/10.1016/S1470-2045(22)00197-8
|
| [75] |
Overman MJ, McDermott R, Leach JL, et al. (2017) Nivolumab in patients with metastatic DNA mismatch repair-deficient or microsatellite instability-high colorectal cancer (CheckMate 142): an open-label, multicentre, phase 2 study. Lancet Oncol 18: 1182-1191. https://doi.org/10.1016/S1470-2045(17)30422-9
|
| [76] |
Overman MJ, Lonardi S, Wong KYM, et al. (2018) Durable clinical benefit with nivolumab plus ipilimumab in DNA mismatch repair-deficient/microsatellite instability-high metastatic colorectal cancer. J Clin Oncol 36: 773-779. https://doi.org/10.1200/JCO.2017.76.9901
|
| [77] |
Xie YH, Chen YX, Fang JY (2020) Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct Target Ther 5: 22. https://doi.org/10.1038/s41392-020-0116-z
|
| [78] |
Saltz LB, Clarke S, Díaz-Rubio E, et al. (2008) Bevacizumab in combination with oxaliplatin-based chemotherapy as first-line therapy in metastatic colorectal cancer: a randomized phase III study. J Clin Oncol 26: 2013-2019. https://doi.org/10.1200/JCO.2007.14.9930
|
| [79] |
Van Cutsem E, Tabernero J, Lakomy R, et al. (2012) Addition of aflibercept to fluorouracil, leucovorin, and irinotecan improves survival in a phase III randomized trial in patients with metastatic colorectal cancer previously treated with an oxaliplatin-based regimen. J Clin Oncol 30: 3499-3506. https://doi.org/10.1200/JCO.2012.42.8201
|
| [80] | Siravegna G, Mussolin B, Buscarino M, et al. (2015) Clonal evolution and resistance to EGFR blockade in the blood of colorectal cancer patients. Nat Med 21: 795-801. https://doi.org/10.1038/nm.3870 |
| [81] |
Goede V, Coutelle O, Neuneier J, et al. (2010) Identification of serum angiopoietin-2 as a biomarker for clinical outcome of colorectal cancer patients treated with bevacizumab-containing therapy. Br J Cancer 103: 1407-1414. https://doi.org/10.1038/sj.bjc.6605925
|
| [82] |
Derangère V, Fumet JD, Boidot R, et al. (2016) Does bevacizumab impact anti-EGFR therapy efficacy in metastatic colorectal cancer?. Oncotarget 7: 9309-9321. https://doi.org/10.18632/oncotarget.7008
|
| [83] |
André T, Shiu KK, Kim TW, et al. (2025) Pembrolizumab versus chemotherapy in microsatellite instability-high or mismatch repair-deficient metastatic colorectal cancer: 5-year follow-up from the randomized phase III KEYNOTE-177 study. Ann Oncol 36: 277-284. https://doi.org/10.1016/j.annonc.2024.11.012
|
| [84] |
Shan J, Han D, Shen C, et al. (2022) Mechanism and strategies of immunotherapy resistance in colorectal cancer. Front Immunol 13: 1016646. https://doi.org/10.3389/fimmu.2022.1016646
|
| [85] |
Lenz HJ, Van Cutsem E, Luisa Limon M, et al. (2022) First-line nivolumab plus low-dose ipilimumab for microsatellite instability-high/mismatch repair-deficient metastatic colorectal cancer: The phase II CheckMate 142 study. J Clin Oncol 40: 161-170. https://doi.org/10.1200/JCO.21.01015
|
| [86] |
André T, Lonardi S, Wong KYM, et al. (2022) Nivolumab plus low-dose ipilimumab in previously treated patients with microsatellite instability-high/mismatch repair-deficient metastatic colorectal cancer: 4-year follow-up from CheckMate 142. Ann Oncol 33: 1052-1060. https://doi.org/10.1016/j.annonc.2022.06.008
|
| [87] |
Overman MJ, Gelsomino F, Aglietta M, et al. (2024) Nivolumab plus relatlimab in patients with previously treated microsatellite instability-high/mismatch repair-deficient metastatic colorectal cancer: the phase II CheckMate 142 study. J Immunother Cancer 12: e008689. https://doi.org/10.1136/jitc-2023-008689
|
| [88] |
Lentz RW, Friedrich TJ, Blatchford PJ, et al. (2024) A phase II study of potentiation of pembrolizumab with binimetinib and bevacizumab in refractory microsatellite-stable colorectal cancer. Clin Cancer Res 30: 3768-3778. https://doi.org/10.1158/1078-0432.CCR-24-0090
|
| [89] |
Poto R, Troiani T, Criscuolo G, et al. (2022) Holistic approach to immune checkpoint inhibitor-related adverse events. Front Immunol 13: 804597. https://doi.org/10.3389/fimmu.2022.804597
|
| [90] |
Cunningham M, Gupta R, Butler M (2024) Checkpoint inhibitor hepatotoxicity: pathogenesis and management. Hepatology 79: 198-212. https://doi.org/10.1097/HEP.0000000000000045
|
| [91] |
Wang DY, Salem JE, Cohen JV, et al. (2018) Fatal toxic effects associated with immune checkpoint inhibitors: A systematic review and Meta-analysis. JAMA Oncol 4: 1721-1728. https://doi.org/10.1001/jamaoncol.2018.3923
|
| [92] |
Forbes N, Hilsden RJ, Martel M, et al. (2021) Association between time to colonoscopy after positive fecal testing and colorectal cancer outcomes: A systematic review. Clin Gastroenterol Hepatol 19: 1344-1354.e8. https://doi.org/10.1016/j.cgh.2020.09.048
|
| [93] |
Siegel R, DeSantis C, Virgo K, et al. (2012) Cancer treatment and survivorship statistics, 2012. CA Cancer J Clin 62: 220-241. https://doi.org/10.3322/caac.21149
|
Figures(1) / Tables(2)

Derek A. Corica, Dylan M. Schaap, Trenton G. Mayberry, Braydon C. Cowan, Mark R. Wakefield, Yujiang Fang. New depths: exploring the current landscape of colorectal cancer[J]. AIMS Medical Science, 2025, 12(1): 105-123. doi: 10.3934/medsci.2025008







 DownLoad:
DownLoad: