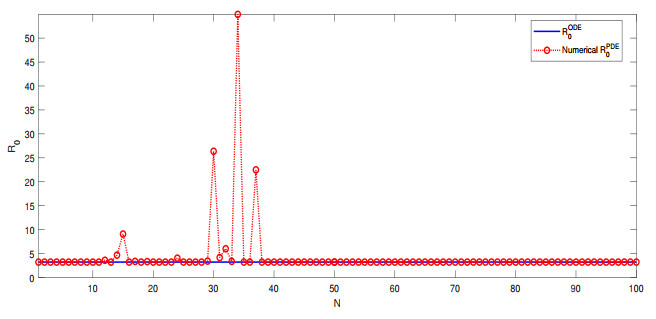
We consider a class of $ k $-dimensional reaction-diffusion epidemic models ($ k = 1, 2, \cdots $) that are developed from autonomous ODE systems. We present a computational approach for the calculation and analysis of their basic reproduction numbers. Particularly, we apply matrix theory to study the relationship between the basic reproduction numbers of the PDE models and those of their underlying ODE models. We show that the basic reproduction numbers are the same for these PDE models and their associated ODE models in several important scenarios. We additionally provide two numerical examples to verify our analytical results.
Citation: Chayu Yang, Jin Wang. Computation of the basic reproduction numbers for reaction-diffusion epidemic models[J]. Mathematical Biosciences and Engineering, 2023, 20(8): 15201-15218. doi: 10.3934/mbe.2023680
We consider a class of $ k $-dimensional reaction-diffusion epidemic models ($ k = 1, 2, \cdots $) that are developed from autonomous ODE systems. We present a computational approach for the calculation and analysis of their basic reproduction numbers. Particularly, we apply matrix theory to study the relationship between the basic reproduction numbers of the PDE models and those of their underlying ODE models. We show that the basic reproduction numbers are the same for these PDE models and their associated ODE models in several important scenarios. We additionally provide two numerical examples to verify our analytical results.

| [1] |
E. Bertuzzo, R. Casagrandi, M. Gatto, I. Rodriguez-Iturbe, A. Rinaldo, On spatially explicit models of cholera epidemics, J. R. Soc. Interface, 7 (2010), 321–333. https://doi.org/10.1098/rsif.2009.0204 doi: 10.1098/rsif.2009.0204

|
| [2] |
R. S. Cantrell, C. Cosner, The effects of spatial heterogeneity in population dynamics, J. Math. Biol., 29 (1991), 315–338. https://doi.org/10.1007/BF00167155 doi: 10.1007/BF00167155
|
| [3] | R. S. Cantrell, C. Cosner, Spatial Ecology via Reaction-Diffusion Equations, Wiley, 2003. https://doi.org/10.1002/0470871296 |
| [4] |
K. I. Kim, Z. Lin, Q. Zhang, An SIR epidemic model with free boundary, Nonlinear Anal. Real World Appl., 14 (2013), 1992–2001. https://doi.org/10.1016/j.nonrwa.2013.02.003 doi: 10.1016/j.nonrwa.2013.02.003
|
| [5] |
A. Rinaldo, E. Bertuzzo, L. Mari, L. Righetto, M. Blokesch, M. Gatto, et al., Reassessment of the 2010-2011 Haiti cholera outbreak and rainfall-driven multiseason projections, PNAS, 109 (2012), 6602–6607. https://doi.org/10.1073/pnas.1203333109 doi: 10.1073/pnas.1203333109
|
| [6] |
F. B. Wang, J. Shi, X. Zou, Dynamics of a host-pathogen system on a bounded spatial domain, Commun. Pure Appl. Anal., 14 (2015), 2535–2560. https://doi.org/10.3934/cpaa.2015.14.2535 doi: 10.3934/cpaa.2015.14.2535
|
| [7] |
X. Wang, D. Gao, J. Wang, Influence of human behavior on cholera dynamics, Math. Biosci., 267 (2015), 41–52. https://doi.org/10.1016/j.mbs.2015.06.009 doi: 10.1016/j.mbs.2015.06.009
|
| [8] |
X. Wang, D. Posny, J. Wang, A reaction-convection-diffusion model for cholera spatial dynamics, Discrete Contin. Dyn. Syst. - Ser. B, 21 (2016), 2785–2809. https://doi.org/10.3934/dcdsb.2016073 doi: 10.3934/dcdsb.2016073
|
| [9] |
O. Diekmann, J. A. P. Heesterbeek, A. J. Metz, On the definition and the computation of the basic reproduction ratio $R_0$ in models for infectious diseases in heterogeneous population, J. Math. Biol., 28 (1990), 365–382. https://doi.org/10.1007/BF00178324 doi: 10.1007/BF00178324
|
| [10] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [11] |
H. R. Thieme, Spectral bound and reproduction number for infinite-dimensional population structure and time heterogeneity, SIAM J. Appl. Math., 70 (2009), 188–211. https://doi.org/10.1137/080732870 doi: 10.1137/080732870
|
| [12] |
W. Wang, X. Q. Zhao, Basic reproduction numbers for reaction-diffusion epidemic models, SIAM J. Appl. Dyn. Syst., 11 (2012), 1652–1673. https://doi.org/10.1137/120872942 doi: 10.1137/120872942
|
| [13] |
L. J. S. Allen, B. M. Bolker, Y. Lou, A. L. Nevai, Asymptotic profiles of the steady states for an SIS epidemic reaction-diffusion model, Discrete Contin. Dyn. Syst., 21 (2008), 1–20. https://doi.org/10.3934/dcds.2008.21.1 doi: 10.3934/dcds.2008.21.1
|
| [14] |
S. Chen, J. Shi, Asymptotic profiles of basic reproduction number for epidemic spreading in heterogeneous environment, SIAM J. Appl. Math., 80 (2020), 1247–1271. https://doi.org/10.1137/19M128907 doi: 10.1137/19M128907
|
| [15] |
P. Magal, G. F. Webb, Y. Wu, On the basic reproduction number of reaction-diffusion epidemic models, SIAM J. Appl. Math., 79 (2019), 284–304. https://doi.org/10.1137/18M1182243 doi: 10.1137/18M1182243
|
| [16] |
P. Song, Y. Lou, Y. Xiao, A spatial SEIRS reaction-diffusion model in heterogeneous environment, J. Differ. Equations, 267 (2019), 5084–5114. https://doi.org/10.1016/j.jde.2019.05.022 doi: 10.1016/j.jde.2019.05.022
|
| [17] |
X. Lin, Q. Wang, Asymptotic behavior of the principal eigenvalue and basic reproduction ratio for time-periodic reaction-diffusion systems with time delay, Discrete Contin. Dyn. Syst. - Ser. B, 28 (2023), 3955–3984. https://doi.org/0.3934/dcdsb.2022250 doi: 10.3934/dcdsb.2022250
|
| [18] |
L. Zhang, X. Q. Zhao, Asymptotic behavior of the basic reproduction ratio for periodic reaction-diffusion systems, SIAM J. Math. Anal., 53 (2021), 6873–6909. https://doi.org/10.1137/20M1366344 doi: 10.1137/20M1366344
|
| [19] |
C. Barril, A. Calsina, J. Ripoll, A practical approach to R0 in continuous-time ecological models, Math. Methods Appl. Sci., 41 (2018), 8432–8445. https://doi.org/10.1002/mma.4673 doi: 10.1002/mma.4673
|
| [20] |
D. Breda, F. Florian, J. Ripoll, R. Vermiglio, Efficient numerical computation of the basic reproduction number for structured populations, J. Comput. Appl. Math., 384 (2021), 113165. https://doi.org/10.1016/j.cam.2020.113165 doi: 10.1016/j.cam.2020.113165
|
| [21] |
J. Ge, C. Lei, Z. Lin, Reproduction numbers and the expanding fronts for a diffusion-advection SIS model in heterogeneous time-periodic environment, Nonlinear Anal. Real World Appl., 33 (2017), 100–120. https://doi.org/10.1016/j.nonrwa.2016.06.005 doi: 10.1016/j.nonrwa.2016.06.005
|
| [22] |
Y. Lou, X. Q. Zhao, A reaction-diffusion malaria model with incubation period in the vector population, J. Math. Biol., 62 (2011), 543–568. https://doi.org/10.1007/s00285-010-0346-8 doi: 10.1007/s00285-010-0346-8
|
| [23] |
H. Zhao, K. Wang, H. Wang, Basic reproduction ratio of a mosquito-borne disease in heterogeneous environment, J. Math. Biol., 86 (2023), 32. https://doi.org/10.1007/s00285-023-01867-y doi: 10.1007/s00285-023-01867-y
|
| [24] |
C. Yang, J. Wang, Basic reproduction numbers for a class of reaction-diffusion epidemic models, Bull. Math. Biol., 82 (2020), 111. https://doi.org/10.1007/s11538-020-00788-x doi: 10.1007/s11538-020-00788-x
|
| [25] | R. D. Richtmyer, K. W. Morton, Difference Methods for Initial-Value Problems, Second Edition, Krieger Publication Company, 1994. |
| [26] | J. W. Thomas, Numerical Partial Differential Equations: Finite Difference Methods, Springer-Verlag New York, 1995. https://doi.org/10.1007/978-1-4899-7278-1 |
| [27] | G. H. Golub, C. F. van Loan, Matrix Computations, Third Edition, Johns Hopkins University Press, 1996. |
| [28] | Y. Saad, Numerical Methods for Large Eigenvalue Problems, Revised Edition, SIAM, 2011. |
| [29] |
C. P. Gerba, Environmentally transmitted pathogens, Environ. Microbiol., 2015 (2015), 509–550. https://doi.org/10.1016/B978-0-12-394626-3.00022-3 doi: 10.1016/B978-0-12-394626-3.00022-3
|
| [30] |
Z. Mukandavire, S. Liao, J. Wang, H. Gaff, D. L. Smith, J. G. Morris, Estimating the reproductive numbers for the 2008-2009 cholera outbreaks in Zimbabwe, PNAS, 108 (2011), 8767–8772. https://doi.org/10.1073/pnas.1019712108 doi: 10.1073/pnas.1019712108
|
| [31] |
D. Posny, J. Wang, Modeling cholera in periodic environments, J. Biol. Dyn., 8 (2014), 1–19. https://doi.org/10.1080/17513758.2014.896482 doi: 10.1080/17513758.2014.896482
|
| [32] |
J. H. Tien, D. J. Earn, Multiple transmission pathways and disease dynamics in a waterborne pathogen model, Bull. Math. Biol., 72 (2010), 1506–1533. https://doi.org/10.1007/s11538-010-9507-6 doi: 10.1007/s11538-010-9507-6
|
| [33] |
T. M. Nguyen, D. Ilef, S. Jarraud, L. Rouil, C. Campese, D. Che, et al., A community-wide outbreak of legionnaires disease linked to industrial cooling towers–How far can contaminated aerosols spread? J. Infect. Dis., 193 (2006), 102–111. https://doi.org/10.1086/498575 doi: 10.1086/498575
|
| [34] |
T. Greenhalgh, J. L. Jimenez, K. A. Prather, Z. Tufekci, D. Fisman, R. Schooley, Ten scientific reasons in support of airborne transmission of SARS-CoV-2, Lancet, 397 (2021), 1603–1605. https://doi.org/10.1016/S0140-6736(21)00869-2 doi: 10.1016/S0140-6736(21)00869-2
|
| [35] |
L. D. Stetzenbach, Airborne infectious microorganisms, Encycl. Microbiol., 2009 (2009), 175–182. https://doi.org/10.1016/B978-012373944-5.00177-2 doi: 10.1016/B978-012373944-5.00177-2
|
| [36] |
K. Yamazaki, C. Yang, J. Wang, A partially diffusive cholera model based on a general second-order differential operator, J. Math. Anal. Appl., 501 (2021), 125181. https://doi.org/10.1016/j.jmaa.2021.125181 doi: 10.1016/j.jmaa.2021.125181
|



Figures(4)

Chayu Yang, Jin Wang. Computation of the basic reproduction numbers for reaction-diffusion epidemic models[J]. Mathematical Biosciences and Engineering, 2023, 20(8): 15201-15218. doi: 10.3934/mbe.2023680







 DownLoad:
DownLoad: