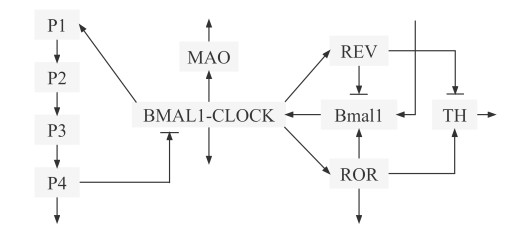
The circadian clock is an autonomous timing system that regulates the physiological and behavioral activities of organisms. Dopamine (DA) is an important neurotransmitter that is associated with many biological activities such as mood and movement. Experimental studies have shown that the circadian clock influences the DA system and disorders in the circadian clock lead to DA-related diseases. However, the regulatory mechanism of the circadian clock on DA is far from clear. In this paper, we apply an existing circadian-dopamine mathematical model to explore the effects of the circadian clock on DA. Based on numerical simulations, we find the disturbance of the circadian clock, including clock gene mutations, jet lag and light pulses, leads to abnormal DA levels. The effects of mutations in some clock genes on the mood and behavior of mice are closely related to DA disruptions. By sensitivity analysis of DA levels to parameter perturbation, we identify key reactions that affect DA levels, which provides insights into modulating DA disorders. Sudden changes in external light influence the circadian clock, bringing about effects on the DA system. Jet lag causes transient DA rhythm desynchronization with the environment and the influence of jet lag in different directions on DA level and phase varies. Light pulses affect the amplitude and phase shift of DA, which provides a promising method for treating DA disorders through light exposure. This study helps to better understand the impact of the circadian clock on the DA system and provides theoretical support for the treatment of DA disorders.
Citation: Ying Li, Zhao Zhao, Yuan-yuan Tan, Xue Wang. Dynamical analysis of the effects of circadian clock on the neurotransmitter dopamine[J]. Mathematical Biosciences and Engineering, 2023, 20(9): 16663-16677. doi: 10.3934/mbe.2023742
The circadian clock is an autonomous timing system that regulates the physiological and behavioral activities of organisms. Dopamine (DA) is an important neurotransmitter that is associated with many biological activities such as mood and movement. Experimental studies have shown that the circadian clock influences the DA system and disorders in the circadian clock lead to DA-related diseases. However, the regulatory mechanism of the circadian clock on DA is far from clear. In this paper, we apply an existing circadian-dopamine mathematical model to explore the effects of the circadian clock on DA. Based on numerical simulations, we find the disturbance of the circadian clock, including clock gene mutations, jet lag and light pulses, leads to abnormal DA levels. The effects of mutations in some clock genes on the mood and behavior of mice are closely related to DA disruptions. By sensitivity analysis of DA levels to parameter perturbation, we identify key reactions that affect DA levels, which provides insights into modulating DA disorders. Sudden changes in external light influence the circadian clock, bringing about effects on the DA system. Jet lag causes transient DA rhythm desynchronization with the environment and the influence of jet lag in different directions on DA level and phase varies. Light pulses affect the amplitude and phase shift of DA, which provides a promising method for treating DA disorders through light exposure. This study helps to better understand the impact of the circadian clock on the DA system and provides theoretical support for the treatment of DA disorders.

| [1] |
J. D. Berke, What does dopamine mean?, Nature Neurosci., 21 (2018), 787. https://doi.org/10.1038/s41593-018-0152-y doi: 10.1038/s41593-018-0152-y

|
| [2] |
A. M. Young, M. H. Joseph, J. A. Gray, Increased dopamine release in vivo in nucleus accumbens and caudate nucleus of the rat during drinking: a microdialysis study, Neuroscience, 48 (1992), 871. https://doi.org/10.2307/941706 doi: 10.2307/941706
|
| [3] |
A. L. Brody, M. A. Mandelkern, R. E. Olmstead, D. Scheibal, E. Hahn, S. Shiraga, et al., Gene variants of brain dopamine pathways and smoking-induced dopamine release in the ventral caudate/nucleus accumbens, Arch. Gen. Psychiatry, 63 (2006), 808. https://doi.org/10.1001/archpsyc.63.7.714 doi: 10.1001/archpsyc.63.7.714
|
| [4] |
M. A. Sacheli, J. L. Neva, B. Lakhani, D. K. Murray, N. Vafai, E. Shahinfard, et al., Exercise increases caudate dopamine release and ventral striatal activation in Parkinson's disease, Mov. Disord., 34 (1891). https://doi.org/10.1002/mds.27865 doi: 10.1002/mds.27865
|
| [5] |
B. Radwan, H. Liu, D. Chaudhury, The role of dopamine in mood disorders and the associated changes in circadian rhythms and sleep-wake cycle, Brain Res., 1713 (2019), 42. https://doi.org/10.1016/j.brainres.2018.11.031 doi: 10.1016/j.brainres.2018.11.031
|
| [6] |
M. Hastings, E. S. Maywood, Circadian clocks in the mammalian brain, Bioessays, 22 (2000), 23. https://doi.org/10.1002/(SICI)1521-1878(200001)22:1<23::AID-BIES6>3.0.CO;2-Z doi: 10.1002/(SICI)1521-1878(200001)22:1<23::AID-BIES6>3.0.CO;2-Z
|
| [7] |
P. L. Lowrey, J. S. Takahashi, Mammalian circadian biology: elucidating genome-wide levels of temporal organization, Annu. Rev. Genomics Hum. Genet., 5 (2004), 407. https://doi.org/10.1146/annurev.genom.5.061903.175925 doi: 10.1146/annurev.genom.5.061903.175925
|
| [8] |
S. A. Brown, G. Zumbrunn, F. Fleury-Olela, N. Preitner, U. Schibler, Rhythms of mammalian body temperature can sustain peripheral circadian clocks, Current Biol., 12 (2002), 1574. https://doi.org/10.1016/S0960-9822(02)01145-4 doi: 10.1016/S0960-9822(02)01145-4
|
| [9] |
A. L. Baird, A. N. Coogan, A. Siddiqui, R. M. Donev, J. Thome, Adult attention-deficit hyperactivity disorder is associated with alterations in circadian rhythms at the behavioural, endocrine and molecular levels, Mol. Psychiatry, 17 (2012), 988. https://doi.org/10.1038/mp.2011.149 doi: 10.1038/mp.2011.149
|
| [10] | J. Huang, Z. M. Zhong, M. Y. Wang, X. F. Chen, Y. C. Tan, S. Q. Zhang, et al., Circadian modulation of dopamine levels and dopaminergic neuron development contributes to attention deficiency and hyperactive behavior, J. Neurosci., 35 (2015), 2572. https://www.jneurosci.org/content/35/6/2572 |
| [11] |
R. Kim, M. C. Reed, A mathematical model of circadian rhythms and dopamine, Theor. Biol. Med. Model., 18 (2021), 8. https://doi.org/10.1186/s12976-021-00139-w doi: 10.1186/s12976-021-00139-w
|
| [12] |
R. Kim, T. P. Witelski, Uncovering the dynamics of a circadian-dopamine model influenced by the light-dark cycle, Math. Biosci., 344 (2022), 108764. https://doi.org/10.1016/j.mbs.2021.108764 doi: 10.1016/j.mbs.2021.108764
|
| [13] |
L. Gravotta, A. M. Gavrila, S. Hood, S. Amir, Global depletion of dopamine using intracerebroventricular 6-hydroxydopamine injection disrupts normal circadian wheel-running patterns and period2 expression in the rat forebrain, J. Mol. Neurosci., 45 (2011), 162. https://doi.org/10.1007/s12031-011-9520-8 doi: 10.1007/s12031-011-9520-8
|
| [14] |
M. Imbesi, S. Yildiz, A. D. Arslan, R. Sharma, H. Manev, T. Uz, Dopamine receptor-mediated regulation of neuronal clock gene expression, Neuroscience, 158 (2009), 537. https://doi.org/10.1016/j.neuroscience.2008.10.044 doi: 10.1016/j.neuroscience.2008.10.044
|
| [15] |
G. Hampp, J. A. Ripperger, T. Houben, I. Schmutz, C. Blex, S. Perreau-Lenz, et al., Regulation of monoamine oxidase A by circadian-clock components implies clock influence on mood, Curr. Biol., 18 (2008), 678. https://doi.org/10.1016/j.cub.2008.04.012 doi: 10.1016/j.cub.2008.04.012
|
| [16] |
J. C. Leloup, A. Goldbeter, Toward a detailed computational model for the mammalian circadian clock, Proc. Natl. Acad. Sci. U. S. A., 100 (2003), 7051. https://doi.org/10.1073/pnas.1132112100 doi: 10.1073/pnas.1132112100
|
| [17] |
Y. Li, G. K. Zhang, Z. G. Song, Entrainment mechanism of the cyanobacterial circadian clock induced by oxidized quinone, Chin. Phys. B, 29 (2020). https://doi.org/10.1088/1674-1056/aba615 doi: 10.1088/1674-1056/aba615
|
| [18] |
Y. Li, G. K. Zhang, Y. M. Ge, Dynamic behavior of the cyanobacterial circadian clock with regulation of CikA, Chin. Phys. B, 30 (2021), 108702. https://doi.org/10.1088/1674-1056/abfb54 doi: 10.1088/1674-1056/abfb54
|
| [19] |
J. A. Best, H. F. Nijhout, M. C. Reed, Homeostatic mechanisms in dopamine synthesis and release: a mathematical model, Theor. Biol. Med. Model., 6 (2009). https://doi.org/10.1186/1742-4682-6-21 doi: 10.1186/1742-4682-6-21
|
| [20] |
C. S. Colwell, Linking neural activity and molecular oscillations in the SCN, Nat. Rev. Neurosci., 12 (2015), 553. https://doi.org/10.1038/nrn3086 doi: 10.1038/nrn3086
|
| [21] |
M. Dadon-Freiberg, N. Chapnik, O. Froy, REV-ERB$\alpha$ alters circadian rhythms by modulating mTOR signaling, Nat. Rev. Neurosci., 521 (2021), 111108. https://doi.org/10.1016/j.mce.2020.111108 doi: 10.1016/j.mce.2020.111108
|
| [22] |
M. Akashi, T. Takumi, The orphan nuclear receptor RORalpha regulates circadian transcription of the mammalian core-clock Bmal1, Nat. Struct. Mol. Biol., 12 (2005), 441. https://doi.org/10.1038/nsmb925 doi: 10.1038/nsmb925
|
| [23] |
S. Chung, E. J. Lee, S. Yun, H. K. Choe, S. B. Park, H. J. Son, et al., Impact of circadian nuclear receptor REV-ERB$\alpha$ on midbrain dopamine production and mood regulation, Cell, 157 (2014), 858. https://doi.org/10.1016/j.cell.2014.03.039 doi: 10.1016/j.cell.2014.03.039
|
| [24] |
M. H. Vitaterna, C. H. Ko, A. M. Chang, E. D. Buhr, E. M. Fruechte, A. Schook, et al., The mouse clock mutation reduces circadian pacemaker amplitude and enhances efficacy of resetting stimuli and phase-response curve amplitude, Cell, 103 (2006), 9327. https://doi.org/10.1073/pnas.0603601103 doi: 10.1073/pnas.0603601103
|
| [25] |
M. Kohiyama, D. Bonser, L. Leung, A. Fall, N. Canada, B. Y. Qu, et al., The Drosophila apterous (56f) mutation impairs circadian locomotor activity, Biol. Rhythm Res., 50 (2018), 375. https://doi.org/10.1080/09291016.2018.1447353 doi: 10.1080/09291016.2018.1447353
|
| [26] |
S. Langmesser, T. Tallone, A. Bordon, S. Rusconi, U. Albrecht, Interaction of circadian clock proteins PER2 and CRY with BMAL1 and CLOCK, BMC Mol. Biol., 9 (2008). https://doi.org/10.1186/1471-2199-9-41 doi: 10.1186/1471-2199-9-41
|
| [27] |
C. A. McClung, K. Sidiropoulou, M. Vitaterna, J. S. Takahashi, F. J. White, D. C. Cooper, et al., Regulation of dopaminergic transmission and cocaine reward by the Clock gene, Proc. Natl. Acad. Sci. U. S. A., 102 (2005), 9377. https://doi.org/10.1073/pnas.0503584102 doi: 10.1073/pnas.0503584102
|
| [28] |
M. M. Sidor, S. M. Spencer, K. Dzirasa, P. K. Parekh, K. M. Tye, M. R. Warden, et al., Daytime spikes in dopaminergic activity drive rapid mood-cycling in mice, Mol. Psychiatr., 20 (2015), 1479. https://doi.org/10.1038/mp.2014.167 doi: 10.1038/mp.2014.167
|
| [29] |
N. de Zavalia, K. Schoettner, J. A. Goldsmith, P. Solis, S. Ferraro, G. Parent, Bmal1 in the striatum influences alcohol intake in a sexually dimorphic manner, Commun. Biol., 4 (2021), 1227. https://doi.org/10.1038/s42003-021-02715-9 doi: 10.1038/s42003-021-02715-9
|
| [30] |
K. H. Price, H. Dziema, S. Aten, J. Loeser, F. E. Norona, K. Hoyt, et al., Modulation of learning and memory by the targeted deletion of the circadian clock gene Bmal1 in forebrain circuits, Behav. Brain Res., 308 (2016), 222. https://doi.org/10.1016/j.bbr.2016.04.027 doi: 10.1016/j.bbr.2016.04.027
|
| [31] |
K. Schoettner, M. Alonso, M. Button, C. Goldfarb, J. Herrera, N. Quteishat, et al., Characterization of affective behaviors and motor functions in mice with a striatal-specific deletion of bmal1 and per2, Front. Physiol., 13 (2022). https://doi.org/10.3389/fphys.2022.922080/full doi: 10.3389/fphys.2022.922080/full
|
| [32] | J. Aschoff, K. Hoffmann, R. Wever, Re-entrainment of circadian rhythms after phase-shifts of the Zeitgeber, Chronobiologia, 2 (1975), 23. https://pubmed.ncbi.nlm.nih.gov/1192905/ |
| [33] |
G. Katz, R. Durst, Y. Zislin, Y. Barel, H. Y. Knobler, Psychiatric aspects of jet lag: review and hypothesis, Med. Hypotheses, 56 (2001), 20. https://doi.org/10.1054/mehy.2000.1094 doi: 10.1054/mehy.2000.1094
|
| [34] |
J. Arendt, B. Stone, D. J. Skene, Sleep disruption in jet lag and other circadian rhythm-related disorders, Princ. Pract. Sleep Med., (2005), 659. https://doi.org/10.1016/B0-72-160797-7/50062-8 doi: 10.1016/B0-72-160797-7/50062-8
|
| [35] |
A. Herxheimer, J. Waterhouse, The prevention and treatment of jet lag, Br. Med. J., 326 (2003), 296. https://doi.org/10.1136/bmj.326.7384.296 doi: 10.1136/bmj.326.7384.296
|
| [36] |
J. Brainard, M. Gobel, B. Scott, M. Koeppen, T. Eckle, Health implications of disrupted circadian rhythms and the potential for daylight as therapy, Anesthesiology, 122 (2015), 1170. https://doi.org/10.1097/ALN.0000000000000596 doi: 10.1097/ALN.0000000000000596
|
| [37] |
E. Cawley, M. Tippler, N. J. Coupland, C. Benkelfat, D. B. Boivin, M. aan het Rot, et al., Dopamine and light: effects on facial emotion recognition, J. Psychopharmacol., 31 (2018), 1225. https://doi.org/10.1177/0269881117711707 doi: 10.1177/0269881117711707
|
| [38] |
T. Endo, R. Matsumura, I. T. Tokuda, T. Yoshikawa, Y. Shigeyoshi, K. Node, et al., Bright light improves sleep in patients with Parkinson's disease: possible role of circadian restoration, Sci. Rep., 10 (2020), 7982. https://doi.org/10.1038/s41598-020-64645-6 doi: 10.1038/s41598-020-64645-6
|
| [39] |
H. Yamadera, T. Ito, H. Suzuki, K. Asayama, R. Ito, S. Endo, Effects of bright light on cognitive and sleep-wake (circadian) rhythm disturbances in Alzheimertype dementia, Psychiatry Clin. Neurosci., 54 (2000). https://doi.org/10.1046/j.1440-1819.2000.00711.x doi: 10.1046/j.1440-1819.2000.00711.x
|
| [40] |
J. Itzhacki, D. Clesse, Y. Goumon, E. J. Van Someren, J. Mendoza, Light rescues circadian behavior and brain dopamine abnormalities in diurnal rodents exposed to a winter-like photoperiod, Brain. Struct. Funct., 223 (2018), 2641. https://doi.org/10.1007/s00429-018-1655-8 doi: 10.1007/s00429-018-1655-8
|





Figures(6)

Ying Li, Zhao Zhao, Yuan-yuan Tan, Xue Wang. Dynamical analysis of the effects of circadian clock on the neurotransmitter dopamine[J]. Mathematical Biosciences and Engineering, 2023, 20(9): 16663-16677. doi: 10.3934/mbe.2023742







 DownLoad:
DownLoad: