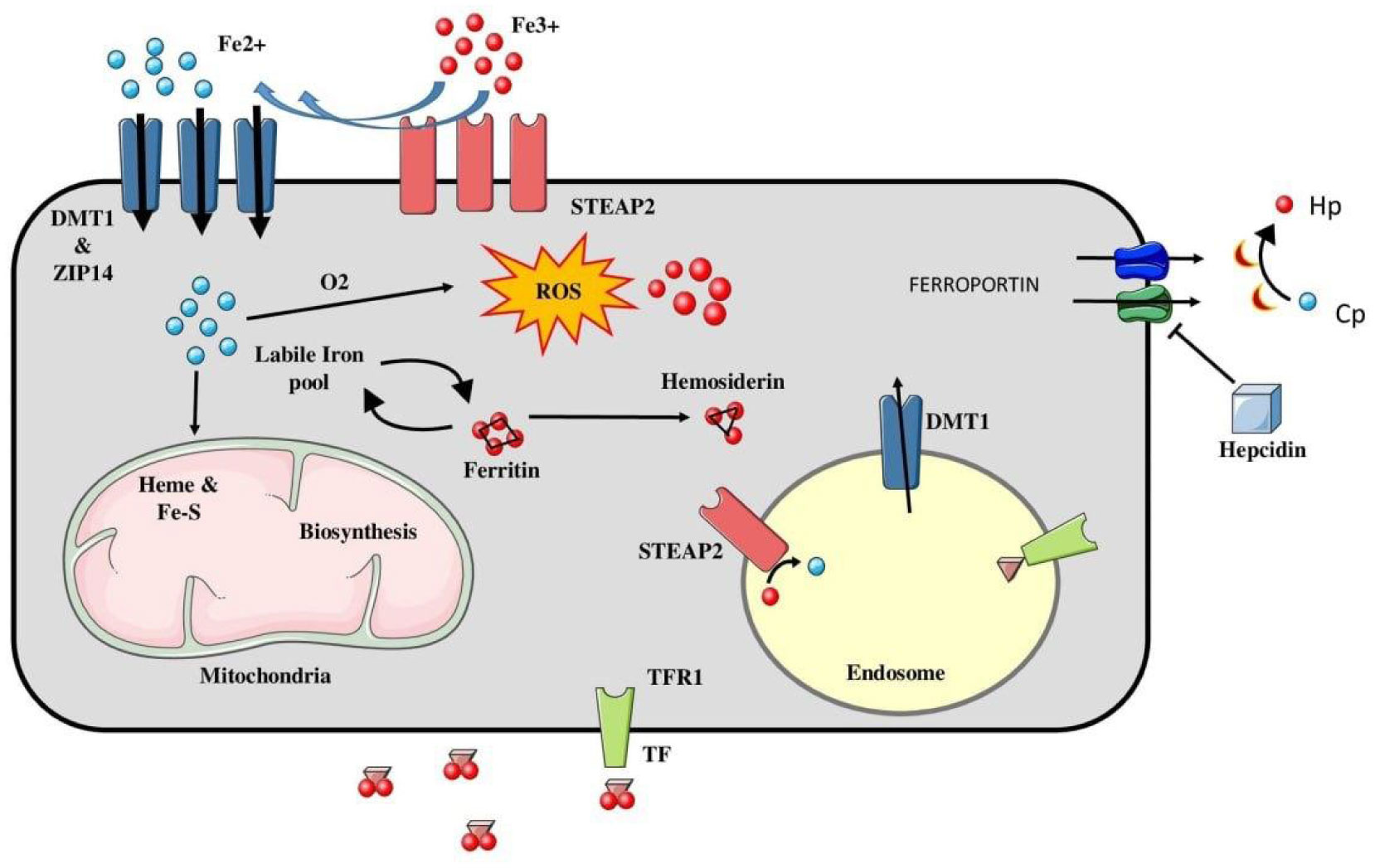
Ferroptosis is a recently discovered iron dependent form of programmed cell death, characterized by accumulation of lipid reactive oxygen species (ROS). It shows a strikingly different set of morphological characteristics from other forms of cell death, like reduced mitochondrial volume, increased bi-layer membrane density, and reduction of mitochondrial cristae with absence of any nuclear changes. Ferroptosis is mainly regulated by two core biochemical processes, namely iron accumulation and lipid peroxidation. Lipid peroxides exert their toxic effects by disturbing the integrity, structure and composition of bi-lipid cell membranes. However, being highly reactive compounds, they further propagate the generation of ROS, leading to cross-linking of DNA and proteins. Key regulators of ferroptosis include various genes involved in the above pathways, inhibition of the antioxidant system and upregulation of the oxidant system. Recent studies have shown the ferroptotic pathway to be involved in the patho-physiology of many diseases, including cancer. Understanding the biochemical mechanisms and key substances upregulating/inhibiting this pathway, may have an implication towards development of targeted therapies for various cancers, and, hence, has become a hotspot for biomedical research. This review article summarizes the core biochemical processes involved in ferroptosis, with a brief summary of its role in various diseases and possible therapeutic targets.
Citation: Naba Hasan, Waleem Ahmad, Feroz Alam, Mahboob Hasan. Ferroptosis-molecular mechanisms and newer insights into some diseases[J]. AIMS Molecular Science, 2023, 10(1): 22-36. doi: 10.3934/molsci.2023003
Ferroptosis is a recently discovered iron dependent form of programmed cell death, characterized by accumulation of lipid reactive oxygen species (ROS). It shows a strikingly different set of morphological characteristics from other forms of cell death, like reduced mitochondrial volume, increased bi-layer membrane density, and reduction of mitochondrial cristae with absence of any nuclear changes. Ferroptosis is mainly regulated by two core biochemical processes, namely iron accumulation and lipid peroxidation. Lipid peroxides exert their toxic effects by disturbing the integrity, structure and composition of bi-lipid cell membranes. However, being highly reactive compounds, they further propagate the generation of ROS, leading to cross-linking of DNA and proteins. Key regulators of ferroptosis include various genes involved in the above pathways, inhibition of the antioxidant system and upregulation of the oxidant system. Recent studies have shown the ferroptotic pathway to be involved in the patho-physiology of many diseases, including cancer. Understanding the biochemical mechanisms and key substances upregulating/inhibiting this pathway, may have an implication towards development of targeted therapies for various cancers, and, hence, has become a hotspot for biomedical research. This review article summarizes the core biochemical processes involved in ferroptosis, with a brief summary of its role in various diseases and possible therapeutic targets.

| [1] |
D'Arcy M (2019) Cell Death. A review of the major forms of Apoptosis, Necrosis and Autophagy. Cell Biol Int 43: 582-592. https://doi.org/10.1002/cbin.11137 
|
| [2] |
Poon IK, Lucas CD, Rossi AG, et al. (2014) Apoptotic cell clearance: basic biology and therapeutic potential. Nat Rev Immunol 14: 166-180. https://doi.org/10.1038/nri3607
|
| [3] |
Bergsbaken T, Fink SL, Cookson BT (2009) Pyroptosis: host cell death and inflammation. Nat Rev Microbiol 7: 99-109. https://doi.org/10.1038/nrmicro2070
|
| [4] |
Yu H, Guo P, Xie X, Wang Y, et al. (2017) Ferroptosis, a new form of cell death, and its relationships with tumourous diseases. J Cell Mol Med 21: 648-657. https://doi.org/10.1111/jcmm.13008
|
| [5] |
Dixon SJ, Lemberg KM, Lamprecht MR, et al. (2012) Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell 149: 1060-1072. https://doi.org/10.1016/j.cell.2012.03.042
|
| [6] |
Kim SE, Zhang L, Ma K, et al. (2016) Ultrasmall nanoparticles induce ferroptosis in nutrient-deprived cancer cells and suppress tumour growth. Nat Nanotechnol 11: 977-985. https://doi.org/10.1038/nnano.2016.164
|
| [7] | Chen X, Yu C, Kang R, et al. (2020) Iron Metabolism in Ferroptosis Front. Cell Dev Biol 8: 590226. http://doi.org/10.3389/fcell.2020.590226 |
| [8] | Stockwell BR., Friedmann Angeli JP, Bayir H, et al. (2017) Ferroptosis: a regulated cell death nexus linking metabolism. Red Biol Dis Cell 171: 273-285. http://doi.org/10.1016/j.cell.2017.09.021 |
| [9] |
Ackermann JA, Hofheinz K, Zaiss MM, et al. (2017) The double-edge role of 12/15-lipoxygenase during inflammation and immunity. Biochim Biophys Acta 862: 371-381. https://doi.org/10.1016/j.bbalip.2016.07.014
|
| [10] |
Yin H, Xu L, Porter NA (2011) Free radical lipid peroxidation: mechanisms and analysis. Chem Rev 111: 5944-5972. https://doi.org/10.1021/cr200084z
|
| [11] |
Tang D, Kroemer G (2020) Ferroptosis. Curr Biol 30: 1292-1297. https://doi.org/10.1016/j.cub.2020.09.068
|
| [12] |
Dolma S, Lessnick SL, Hahn WC, et al. (2003) Identification of genotype-selective antitumor agents using synthetic lethal chemical screening in engineered human tumor cells. Canc Cell 3: 285-296. https://doi.org/10.1016/S1535-6108(03)00050-3
|
| [13] |
Yang WS, Stockwell BR (2008) Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS harboring cancer cells. Chem Biol 15: 234-245. https://doi.org/10.1016/j.chembiol.2008.02.010
|
| [14] |
Yagoda N, Von Rechenberg M, Zaganjor E, et al. (2007) RAS–RAF–MEK-dependent oxidative cell death involving voltage-dependent anion channels. Nat 447: 865-869. https://doi.org/10.1038/nature05859
|
| [15] |
Wenzel SE, Tyurina YY, Zhao J, et al. (2017) PEBP1 wardens ferroptosis by enabling lipoxygenase generation of lipid death signals. Cell 171: 628-641. https://doi.org/10.1016/j.cell.2017.09.044
|
| [16] |
Dai C, Chen X, Li J, et al. (2020) Transcription factors in ferroptotic cell death. Cancer Gene Ther 27: 645-656. https://doi.org/10.1038/s41417-020-0170-2
|
| [17] | Fang X, Ardehali H, Min J, et al. (2022) The molecular and metabolic landscape of iron and ferroptosis in cardiovascular disease. Nat Rev Cardiology 2022: 1-17. https://doi.org/10.1038/s41569-022-00735-4 |
| [18] |
Zhang S, Xin W, Anderson GJ, et al. (2022) Double-edge sword roles of iron in driving energy production versus instigating ferroptosis. Cell Death Dis 13: 40. https://doi.org/10.1038/s41419-021-04490-1
|
| [19] |
Brown CW, Amante JJ, Chhoy P, et al. (2019) Prominin2 drives ferroptosis resistance by stimulating iron export. Dev Cell 51: 575-586. https://doi.org/10.1016/j.devcel.2019.10.007
|
| [20] |
Yang WS, Stockwell BR (2016) Ferroptosis: death by lipid peroxidation. Trends Cell Biol 26: 165-176. https://doi.org/10.1016/j.tcb.2015.10.014
|
| [21] | Yang WS, Kim KJ, Gaschler MM, et al. (2016) Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc Natl Acad Sci USA 113: E4966-E4975. https://doi.org/10.1073/pnas.1603244113 |
| [22] |
Lee H, Zandkarimi F, Zhang Y, et al. (2020) Energy-stress-mediated AMPK activation inhibits ferroptosis. Nat Cell Biol 22: 225-234. https://doi.org/10.1038/s41556-020-0461-8
|
| [23] |
Shin D, Lee J, You JH, et al. (2020) Dihydrolipoamide dehydrogenaseregulates cystine deprivation-induced ferroptosis in head and neck cancer. Redox Biol 30: 101418. https://doi.org/10.1016/j.redox.2019.101418
|
| [24] |
Dixon SJ, Patel DN, Welsch M, et al. (2014) Pharmacological inhibition of cystine–glutamate exchange induces endoplasmic reticulum stress and ferroptosis. elife 3. https://doi.org/10.7554%2FeLife.02523
|
| [25] |
Jiang L, Kon N, Li T, et al. (2015) Ferroptosis as a p53-mediated activity during tumour suppression. Nature 520: 57-62. https://doi.org/10.1038/nature14344
|
| [26] |
Chen D, Tavana O, Chu B, et al. (2017) NRF2 is a major target of ARF in p53-independent tumor suppression. Mol cell 68: 224-232. https://doi.org/10.1016/j.molcel.2017.09.009
|
| [27] |
Zhang Y, Shi J, Liu X, et al. (2018) BAP1 links metabolic regulation of ferroptosis to tumour suppression. Nat Cell Biol 20: 1181-1192. https://doi.org/10.1038/s41556-018-0178-0
|
| [28] |
Yang WS, SriRamaratnam R, Welsch ME, et al. (2014) Regulation of ferroptotic cancer cell death by GPX4. Cell 156: 317-331. https://doi.org/10.1016/j.cell.2013.12.010
|
| [29] |
Bersuker K, Hendricks JM, Li Z, et al. (2019) The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature 575: 688-692. https://doi.org/10.1038/s41586-019-1705-2
|
| [30] |
Shimada K, Hayano M, Pagano NC, et al. (2016) Cell-line selectivity improves the predictive power of pharmacogenomic analyses and helps identify NADPH as biomarker for ferroptosis sensitivity. Cell Chem Biol 23: 225-235. https://doi.org/10.1016/j.chembiol.2015.11.016
|
| [31] |
Jiang L, Hickman JH, Wang SJ, et al. (2015) Dynamic roles of p53-mediated metabolic activities in ROS-induced stress responses. Cell Cycle 14: 2881-2885. https://doi.org/10.1080/15384101.2015.1068479
|
| [32] |
Jiang L, Kon N, Li T, et al. (2015) Ferroptosis as a p53-mediated activity during tumour suppression. Nature 520: 57-62. https://doi.org/10.1038/nature14344
|
| [33] | Ou Y, Wang SJ, Li D, et al. (2016) Activation of SAT1 engages polyamine metabolism with p53-mediated ferroptotic responses. ProcNatl Acad Sci USA 113: 6806-6812. https://doi.org/10.1073/pnas.1607152113 |
| [34] |
Liu J, Zhang C, Wang J, et al. (2020) The Regulation of Ferroptosis by Tumor Suppressor p53 and its Pathway. Int J Mol Sci 21: 8387. https://doi.org/10.3390/ijms21218387
|
| [35] |
Babaei-Abraki S, Karamali F, Nasr-Esfahani MH (2022) Monitoring the induction of ferroptosis following dissociation in human embryonic stem cells. J Biol Chem 298: 101855. https://doi.org/10.1016/j.jbc.2022.101855
|
| [36] |
Nishizawa H, Matsumoto M, Shindo T, et al. (2020) Ferroptosis is controlled by the coordinated transcriptional regulation of glutathione and labile iron metabolism by the transcription factor BACH1. J Biol Chem 295: 69-82. https://doi.org/10.1074/jbc.RA119.009548
|
| [37] |
Wang L, Liu Y, Du T, et al. (2020) ATF3 promotes erastin-induced ferroptosis by suppressing system Xc–. Cell Death Differ 27: 662-675. https://doi.org/10.1038/s41418-019-0380-z
|
| [38] |
Wu J, Minikes AM, Gao M, et al. (2019) Intercellular interaction dictates cancer cell ferroptosis via NF2–YAP signalling. Nature 572: 402-406. https://doi.org/10.1038/s41586-019-1426-6
|
| [39] |
Speer RE, Karuppagounder SS, Basso M, et al. (2013) Hypoxia-inducible factor prolyl hydroxylases as targets for neuroprotection by “antioxidant” metal chelators: From ferroptosis to stroke. Free Radic Biol Med 62: 26-36. https://doi.org/10.1016/j.freeradbiomed.2013.01.026
|
| [40] |
Ou W, Mulik RS, Anwar A, et al. (2017) Low-density lipoprotein docosahexaenoic acid nanoparticles induce ferroptotic cell death in hepatocellular carcinoma. Free Radic Biol Med 112: 597-607. https://doi.org/10.1016/j.freeradbiomed.2017.09.002
|
| [41] |
Louandre C, Marcq I, Bouhlal H, et al. (2015) The retinoblastoma (Rb) protein regulates ferroptosis induced by sorafenib in human hepatocellular carcinoma cells. Cancer Lett 356: 971-977. https://doi.org/10.1016/j.canlet.2014.11.014
|
| [42] |
Bai T, Wang S, Zhao Y, et al. (2017) Haloperidol, a sigma receptor 1 antagonist, promotes ferroptosis in hepatocellular carcinoma cells. Biochem Biophys Res Commun 491: 919-925. https://doi.org/10.1016/j.bbrc.2017.07.136
|
| [43] |
Sun X, Niu X, Chen R, et al. (2016) Metallothionein-1G facilitates sorafenib resistance through inhibition of ferroptosis. Hepatol 64: 488-500. https://doi.org/10.1002/hep.28574
|
| [44] |
Xie Y, Zhu S, Song X, et al. (2017) The tumor suppressor p53 limits ferroptosis by blocking DPP4 activity. Cell Rep 20: 1692-1704. https://doi.org/10.1016/j.celrep.2017.07.055
|
| [45] |
Guo J, Xu B, Han Q, et al. (2018) Ferroptosis: a novel anti-tumor action for cisplatin. Cancer Res Treat 50: 445-460. https://doi.org/10.4143/crt.2016.572
|
| [46] |
Chen MS, Wang SF, Hsu CY, et al. (2017) CHAC1 degradation of glutathione enhances cystine-starvation-induced necroptosis and ferroptosis in human triple negative breast cancer cells via the GCN2-eIF2α-ATF4 pathway. Oncotarget 8: 114588. https://doi.org/10.18632%2Foncotarget.23055
|
| [47] |
Ishimoto T, Nagano O, Yae T, et al. (2011) CD44 variant regulates redox status in cancer cells by stabilizing the xCT subunit of system xc− and thereby promotes tumor growth. Cancer cell 19: 387-400. https://doi.org/10.1016/j.ccr.2011.01.038
|
| [48] | Rui Xiong, Ruyuan He, Bohao Liu, et al. (2021) “Ferroptosis: A New Promising Target for Lung Cancer Therapy”. Oxid Med Cell Longev 2021: 8457521. https://doi.org/10.1155/2021/8457521 |
| [49] |
Zhao H, Xu Y, Shang H (2022) Ferroptosis: A New Promising Target for Ovarian Cancer Therapy. Int J Med Sci 19: 1847-1855. https://doi.org/10.7150/ijms.76480
|
| [50] |
Xu F, Guan Y, Xue L, et al. (2021) The roles of ferroptosis regulatory gene SLC7A11 in renal cell carcinoma: A multi-omics study. Cancer Med 10: 9078-9096.
|
| [51] |
Friedmann Angeli JP, Schneider M, et al. (2014) Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat cell Biol 16: 1180-1191. https://doi.org/10.1038/ncb3064
|
| [52] |
Linkermann A, Skouta R, Himmerkus N, et al. (2014) Synchronized renal tubular cell death involves ferroptosis. Proc Natl Acad Sci USA 111: 16836-16841. https://doi.org/10.1073/pnas.1415518111
|
| [53] |
Martin-Sanchez D, Ruiz-Andres O, Poveda J, et al. (2017) Ferroptosis, but not necroptosis, is important in nephrotoxic folic acid–induced AKI. J Am Soc Nephrol 28: 218-229. https://doi.org/10.1681/ASN.2015121376
|
| [54] |
Li W, Feng G, Gauthier JM, et al. (2019) Ferroptotic cell death and TLR4/Trif signaling initiate neutrophil recruitment after heart transplantation. J clin Investing 129: 2293-2304. https://doi.org/10.1172/JCI126428
|
| [55] |
Fang X, Wang H, Han D, et al. (2019) Ferroptosis as a target for protection against cardiomyopathy. Proc Nationl Acad Sci 116: 2672-2680. https://doi.org/10.1073/pnas.1821022116
|
| [56] |
Dietrich RB, Bradley WG (1988) Iron accumulation in the basal ganglia following severe ischemic-anoxic insults in children. Radiol 168: 203-206. https://doi.org/10.1148/radiology.168.1.3380958
|
| [57] |
Ahmad S, Elsherbiny NM, Haque R, et al. (2014) Sesamin attenuates neurotoxicity in mouse model of ischemic brain stroke. Neurotoxicology 45: 100-110. https://doi.org/10.1016/j.neuro.2014.10.002
|
| [58] |
Hanson LR, Roeytenberg A, Martinez PM, et al. (2009) Intranasal deferoxamine provides increased brain exposure and significant protection in rat ischemic stroke. J Pharmacol Exp Ther 330: 679-686. https://doi.org/10.1124/jpet.108.149807
|
| [59] |
Raven EP, Lu PH, Tishler TA, et al. (2013) Increased iron levels and decreased tissue integrity in hippocampus of Alzheimer's disease detected in vivo with magnetic resonance imaging. J Alzheimers Dis 37: 127-136. http://doi.org/10.3233/JAD-130209
|
| [60] |
Ayton S, Lei P (2014) Nigral iron elevation is an invariable feature of Parkinson's disease and is a sufficient cause of neurodegeneration. BioMed Res Int 2014: 1-9. https://doi.org/10.1155/2014/581256
|
| [61] |
Do Van B, Gouel F, Jonneaux A, et al. (2016) Ferroptosis, a newly characterized form of cell death in Parkinson's disease that is regulated by PKC. Neurobiol Dis 94: 169-178. https://doi.org/10.1016/j.nbd.2016.05.011
|
| [62] |
Klepac N, Relja M, Klepac R, et al. (2007) Oxidative stress parameters in plasma of Huntington's disease patients, asymptomatic Huntington's disease gene carriers and healthy subjects. J Neurol 254: 1676-1683. https://doi.org/10.1007/s00415-007-0611-y
|
| [63] |
Chen J, Marks E, Lai B, et al. (2013) Iron accumulates in Huntington's disease neurons: protection by deferoxamine. PloS one 8: e77023. https://doi.org/10.1371/journal.pone.0077023
|
| [64] |
Kwan JY, Jeong SY, Van Gelderen P, et al. (2012) Iron accumulation in deep cortical layers accounts for MRI signal abnormalities in ALS: correlating 7 tesla MRI and pathology. PloS one 7: e35241. https://doi.org/10.1371/journal.pone.0035241
|
| [65] | Codazzi F, Hu A, Rai M, et al. (2016) Friedreich ataxia-induced pluripotent stem cell-derived neurons show a cellular phenotype that is corrected by a benzamide HDAC inhibitor. Hum Mol Genet 25: 4847-4855. https://doi.org/10.1093/hmg/ddw308 |
| [66] |
Skouta R, Dixon SJ, Wang J, et al. (2014) Ferrostatins inhibit oxidative lipid damage and cell death in diverse disease models. J Am Chem Soc 136: 4551-4556. https://doi.org/10.1021/ja411006a
|
| [67] |
Sun H, Saeedi P, Karuranga S, et al. (2022) IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res Clin Pract 183: 109119. https://doi.org/10.1016/j.diabres.2021.109119
|
| [68] | Wang J, Wang H (2017) Oxidative stress in pancreatic beta cell regeneration. Oxid Med Cell Longev 2017: 1-9. https://doi.org/10.1155/2017/1930261 |
| [69] |
Bruni A, Pepper AR, Pawlick RL, et al. (2018) Ferroptosis-inducing agents compromise in vitro human islet viability and function. Cell Death Dis 9: 595. https://doi.org/10.1038/s41419-018-0506-0
|
| [70] |
Gautam S, Alam F, Moin S, et al. (2021) Role of ferritin and oxidative stress index in gestational diabetes mellitus. J Diabetes Metab Disor 20: 1615-1619. https://doi.org/10.1007/s40200-021-00911-2
|
| [71] |
Lappas M, Hiden U, Desoye G, et al. (2011) The role of oxidative stress in the pathophysiology of gestational diabetes mellitus. Antioxid Redox Signal 15: 3061-3100. https://doi.org/10.1089/ars.2010.3765
|
| [72] |
He J, Li Z, Xia P, et al. (2022) Ferroptosis and ferritinophagy in diabetes complications. Mol Metab 15: 101470. https://doi.org/10.1016/j.molmet.2022.101470
|
| [73] |
Liu Y, Cao X, He C, et al. (2022) Effects of Ferroptosis on Male Reproduction. Int J Mol Sci 23: 7139. https://doi.org/10.3390/ijms23137139
|
| [74] |
Dixon SJ, Lemberg KM, Lamprecht MR, et al. (2012) Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell 149: 1060-1072. https://doi.org/10.1016/j.cell.2012.03.042
|
| [75] |
Shimada K, Skouta R, Kaplan A, et al. (2016) Global survey of cell death mechanisms reveals metabolic regulation of ferroptosis. Nat Chem Biol 12: 497-503. https://doi.org/10.1038/nchembio.2079
|
| [76] |
Abrams RP, Carroll WL, Woerpel KA (2016) Five-membered ring peroxide selectively initiates ferroptosis in cancer cells. ACS Chem Biol 11: 1305-1312. https://doi.org/10.1021/acschembio.5b00900
|
| [77] |
Li C, Zhang Y, Liu J, et al. (2021) Mitochondrial DNA stress triggers autophagy-dependent ferroptotic death. Autophagy 17: 948-960. https://doi.org/10.1080/15548627.2020.1739447
|
| [78] |
Friedmann Angeli JP, Schneider M, et al. (2014) Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nature cell biology 16: 1180-1191. https://doi.org/10.1038/ncb3064
|
| [79] |
Ou M, Jiang Y, Ji Y, et al. (2022) Role and mechanism of ferroptosis in neurological diseases. Mol Metab 61: 101502. https://doi.org/10.1016/j.molmet.2022.101502
|
| [80] | Liu X, Wang T, Wang W, et al. (2022) Emerging Potential Therapeutic Targets of Ferroptosis in Skeletal Diseases. Oxid Med Cell Longev 2022: 3112388. https://doi.org/10.1155/2022/3112388 |


Figures(3)

Naba Hasan, Waleem Ahmad, Feroz Alam, Mahboob Hasan. Ferroptosis-molecular mechanisms and newer insights into some diseases[J]. AIMS Molecular Science, 2023, 10(1): 22-36. doi: 10.3934/molsci.2023003







 DownLoad:
DownLoad: