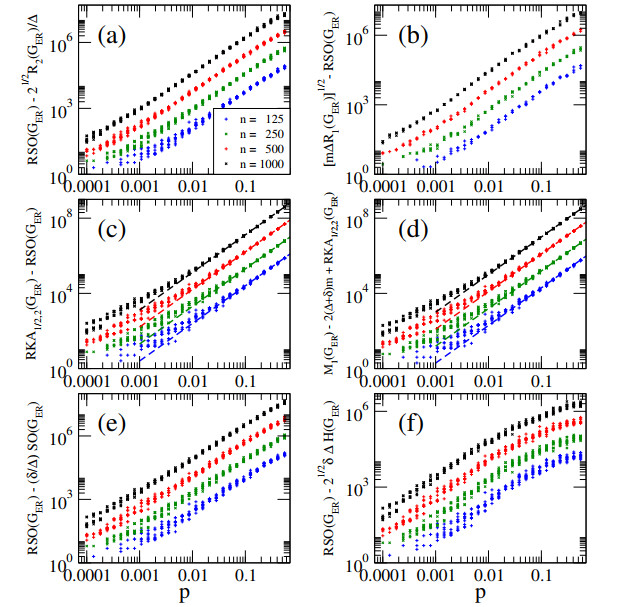
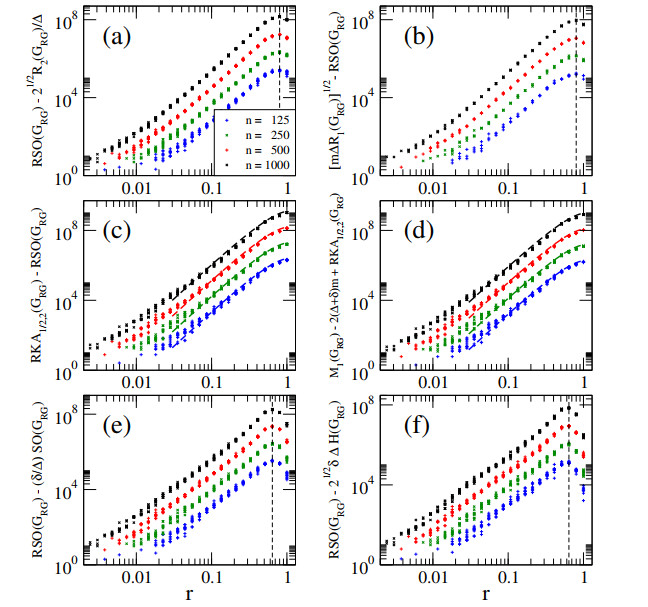
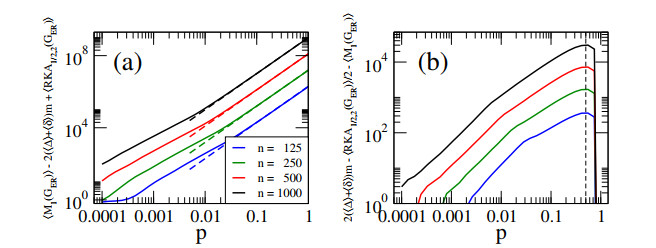
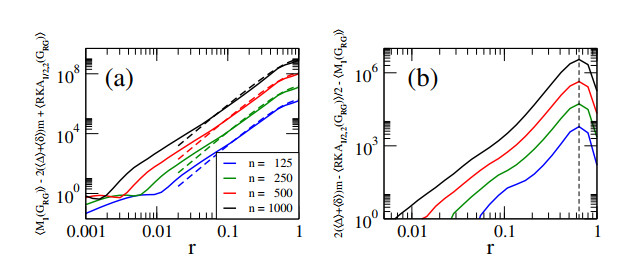
In this paper, we perform analytical and statistical studies of Revan indices on graphs $ G $: $ R(G) = \sum_{uv \in E(G)} F(r_u, r_v) $, where $ uv $ denotes the edge of $ G $ connecting the vertices $ u $ and $ v $, $ r_u $ is the Revan degree of the vertex $ u $, and $ F $ is a function of the Revan vertex degrees. Here, $ r_u = \Delta + \delta - d_u $ with $ \Delta $ and $ \delta $ the maximum and minimum degrees among the vertices of $ G $ and $ d_u $ is the degree of the vertex $ u $. We concentrate on Revan indices of the Sombor family, i.e., the Revan Sombor index and the first and second Revan $ (a, b) $-$ KA $ indices. First, we present new relations to provide bounds on Revan Sombor indices which also relate them with other Revan indices (such as the Revan versions of the first and second Zagreb indices) and with standard degree-based indices (such as the Sombor index, the first and second $ (a, b) $-$ KA $ indices, the first Zagreb index and the Harmonic index). Then, we extend some relations to index average values, so they can be effectively used for the statistical study of ensembles of random graphs.
Citation: V. R. Kulli, J. A. Méndez-Bermúdez, José M. Rodríguez, José M. Sigarreta. Revan Sombor indices: Analytical and statistical study[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 1801-1819. doi: 10.3934/mbe.2023082
In this paper, we perform analytical and statistical studies of Revan indices on graphs $ G $: $ R(G) = \sum_{uv \in E(G)} F(r_u, r_v) $, where $ uv $ denotes the edge of $ G $ connecting the vertices $ u $ and $ v $, $ r_u $ is the Revan degree of the vertex $ u $, and $ F $ is a function of the Revan vertex degrees. Here, $ r_u = \Delta + \delta - d_u $ with $ \Delta $ and $ \delta $ the maximum and minimum degrees among the vertices of $ G $ and $ d_u $ is the degree of the vertex $ u $. We concentrate on Revan indices of the Sombor family, i.e., the Revan Sombor index and the first and second Revan $ (a, b) $-$ KA $ indices. First, we present new relations to provide bounds on Revan Sombor indices which also relate them with other Revan indices (such as the Revan versions of the first and second Zagreb indices) and with standard degree-based indices (such as the Sombor index, the first and second $ (a, b) $-$ KA $ indices, the first Zagreb index and the Harmonic index). Then, we extend some relations to index average values, so they can be effectively used for the statistical study of ensembles of random graphs.

| [1] | I. Gutman, O. E. Polansky, Mathematical Concepts in Organic Chemistry, Springer, Berlin, Heidelberg, 1986. https://doi.org/10.1007/978-3-642-70982-1 |
| [2] | V. R. Kulli, Multiplicative Connectivity Indices of Nanostructures, LAP LEMBERT Academic Publishing, Rīgā, 2018. http://dx.doi.org/10.22147/jusps-A/290101 |
| [3] | V. R. Kulli, College Graph Theory, Vishwa International Publications, Gulbarga, India, 2012. |
| [4] | V. R. Kulli, Revan indices of oxide and honeycomb networks, Int. J. Math. Appl., 55 (2017), 7. |
| [5] | V. R. Kulli, F-Revan index and F-Revan polynomial of some families of benzenoid systems, J. Global Res. Math. Arch., 5 (2018), 1–6. |
| [6] | I. Gutman, Geometric approach to degree based topological MATCH Commun. Math. Comput. Chem., 86 (2021), 11–16. |
| [7] | V. R. Kulli, $ \delta$-Sombor index and its exponential for certain nanotubes, Ann. Pure Appl. Math., 23 (2021), 37–42. |
| [8] | V. R. Kulli, On Banhatti-Sombor indices, SSRG Int. J. Appl. Chem., 8 (2021), 21–25. https://doi.org/10.14445/23939133/IJAC-V8I1P105 |
| [9] |
V. R. Kulli, Sombor indices of certain graph operators, Int. J. Eng. Sci. Res. Technol., 10 (2021), 127–134. https://doi.org/10.29121/ijesrt.v10.i1.2021.12 doi: 10.29121/ijesrt.v10.i1.2021.12

|
| [10] |
V. R. Kulli, I. Gutman, Computation of Sombor indices of certain networks, SSRG Int. J. Appl. Chem., 8 (2021), 1–5. https://doi.org/10.14445/23939133/IJAC-V8I1P101 doi: 10.14445/23939133/IJAC-V8I1P101
|
| [11] |
A. Aashtab, S. Akbari, S. Madadinia, M. Noei, F. Salehi, On the graphs with minimum Sombor index, MATCH Commun. Math. Comput. Chem., 88 (2022), 553–559. https://doi.org/10.46793/match.88-3.553A doi: 10.46793/match.88-3.553A
|
| [12] | B. Horoldagva, C. Xu, On Sombor index of graphs, MATCH Commun. Math. Comput. Chem., 86 (2021), 703–713. |
| [13] |
H. Liu, I. Gutman, L. You, Y. Huang, Sombor index: review of extremal results and bounds, J. Math. Chem., 60 (2022), 771–798. https://doi.org/10.1007/s10910-022-01333-y doi: 10.1007/s10910-022-01333-y
|
| [14] | S. Alikhani, N. Ghanbari, Sombor index of polymers, MATCH Commun. Math. Comput. Chem., 86 (2021), 715–728. |
| [15] |
Y. Shang, Sombor index and degree-related properties of simplicial networks, Appl. Math. Comput., 419 (2022), 126881. https://doi.org/10.1016/j.amc.2021.126881 doi: 10.1016/j.amc.2021.126881
|
| [16] | V. R. Kulli, I. Gutman, Revan Sombor index, J. Math. Inf., 22 (2022), 23–27. |
| [17] |
V. R. Kulli, The $(a, b)$-KA indices of polycyclic aromatic hydrocarbons and benzenoid systems, Int. J. Math. Trends Technol., 65 (2019), 115–120. https://doi.org/10.14445/22315373/IJMTT-V65I11P512 doi: 10.14445/22315373/IJMTT-V65I11P512
|
| [18] | V. R. Kulli, The $(a, b)$-status index of graphs, Ann. Pure Appl. Math., 21 (2020), 113–118. http://dx.doi.org/10.22457/apam.v21n2a5646 |
| [19] |
V. R. Kulli, B. Chaluvaraju, T. Vidya, Computation of Adriatic $(a, b)$-KA index of some nanostructures, Int. J. Math. Trends Technol., 67 (2021), 79–87. 10.14445/22315373/IJMTT-V67I4P511 doi: 10.14445/22315373/IJMTT-V67I4P511
|
| [20] |
V. R. Kulli, I. Gutman, $(a, b)$-KA indices of benzenoid systems and phenylenes: The general case, Int. J. Math. Trends Technol., 67 (2021), 17–20. 10.14445/22315373/IJMTT-V67I1P503 doi: 10.14445/22315373/IJMTT-V67I1P503
|
| [21] | P. P. Korovkin, Inequalities (Little Mathematics Library), Mir Publishers, Moscow, 1975. |
| [22] |
S. Fajtlowicz, On conjectures of Graffiti-II, Ann. Discrete Math., 38 (1988), 113–118. https://doi.org/10.1016/S0167-5060(08)70776-3 doi: 10.1016/S0167-5060(08)70776-3
|
| [23] |
H. Deng, S. Balachandran, S. K. Ayyaswamy, Y. B. Venkatakrishnan, On the harmonic index and the chromatic number of a graph, Discrete Appl. Math., 161 (2013), 2740–2744. https://doi.org/10.1016/j.dam.2013.04.003 doi: 10.1016/j.dam.2013.04.003
|
| [24] |
O. Favaron, M. Mahéo, J. F. Saclé, Some eigenvalue properties in graphs (conjectures of Graffiti-II), Discrete Math., 111 (1993), 197–220. https://doi.org/10.1016/0012-365X(93)90156-N doi: 10.1016/0012-365X(93)90156-N
|
| [25] |
R. Wu, Z. Tanga, H. Deng, A lower bound for the harmonic index of a graph with minimum degree at least two, Filomat, 27 (2013), 51–55. https://doi.org/10.2298/FIL1301051W doi: 10.2298/FIL1301051W
|
| [26] |
L. Zhong, The harmonic index for graphs, Appl. Math. Letters, 25 (2012), 561–566. https://doi.org/10.1016/j.aml.2011.09.059 doi: 10.1016/j.aml.2011.09.059
|
| [27] | L. Zhong, K. Xu, Inequalities between vertex-degree-based topological Indices, MATCH Commun. Math. Comput. Chem., 71 (2014), 627–642. |
| [28] |
R. Aguilar-Sanchez, I. F. Herrera-Gonzalez, J. A. Mendez-Bermudez, J. M. Sigarreta, Computational properties of general indices on random networks, Symmetry, 12 (2020), 1341. https://doi.org/10.3390/sym12081341 doi: 10.3390/sym12081341
|
| [29] |
C. T. Martinez-Martinez, J. A. Mendez-Bermudez, J. M. Rodriguez, J. M. Sigarreta, Computational and analytical studies of the Randić index in Erdös-Rényi models, Appl. Math. Comput., 377 (2020), 125137. https://doi.org/10.1016/j.amc.2020.125137 doi: 10.1016/j.amc.2020.125137
|
| [30] | C. T. Martinez-Martinez, J. A. Mendez-Bermudez, J. M. Rodriguez, J. M. Sigarreta, Computational and analytical studies of the harmonic index in Erdös-Rényi models, MATCH Commun. Math. Comput. Chem., 85 (2021), 395–426. |
| [31] |
R. Aguilar-Sanchez, J. A. Mendez-Bermudez, F. A. Rodrigues, J. M. Sigarreta-Almira, Topological versus spectral properties of random geometric graphs, Phys. Rev. E, 102 (2020), 042306. https://doi.org/10.1103/PhysRevE.102.042306 doi: 10.1103/PhysRevE.102.042306
|
| [32] |
R. Aguilar-Sanchez, J. A. Mendez-Bermudez, J. M. Rodriguez, J. M. Sigarreta, Normalized Sombor indices as complexity measures of random networks, Entropy, 23 (2021), 976. https://doi.org/10.3390/e23080976 doi: 10.3390/e23080976
|
| [33] |
Y. Shang, Estrada index and Laplacian Estrada index of random interdependent graphs, Mathematics, 8 (2020), 1063. https://doi.org/10.3390/math8071063 doi: 10.3390/math8071063
|
| [34] |
R. Aguilar-Sanchez, J. A. Mendez-Bermudez, J. M. Rodriguez, J. M. Sigarreta-Almira, Analytical and statistical studies of Rodriguez–Velazquez indices, J. Math. Chem., 59 (2021), 1246–1259. https://doi.org/10.1007/s10910-021-01239-1 doi: 10.1007/s10910-021-01239-1
|
| [35] | P. Erdös, A. Rényi, On random graphs, Publ. Math. (Debrecen), 6 (1959), 290–297. |
| [36] | R. Solomonoff, A. Rapoport, Connectivity of random nets, Bull. Math. Biophys., 13 (1951), 107–117. https://doi.org/10.1007/BF02478357 |
| [37] | J. Dall, M. Christensen, Random geometric graphs, Phys. Rev. E, 66 (2002), 016121. https://doi.org/10.1103/PhysRevE.66.016121 |
| [38] | M. Penrose, Random Geometric Graphs, Oxford University Press, Oxford, 2003. https://doi.org/10.1093/acprof:oso/9780198506263.001.0001 |
| [39] | R. Aguilar-Sanchez, I. F. Herrera-Gonzalez, J. A. Mendez-Bermudez, J. M. Sigarreta, Revan-degree indices on random graphs, preprint, arXiv: 2210.04749. |
Figures(4)

V. R. Kulli, J. A. Méndez-Bermúdez, José M. Rodríguez, José M. Sigarreta. Revan Sombor indices: Analytical and statistical study[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 1801-1819. doi: 10.3934/mbe.2023082







 DownLoad:
DownLoad: