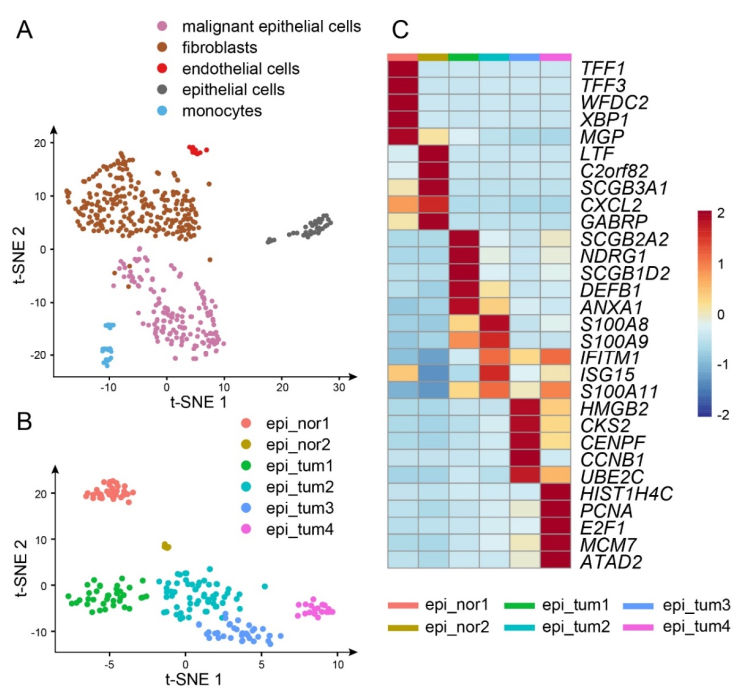
Triple-negative breast cancer (TNBC) is an aggressive subtype of mammary carcinoma characterized by low expression levels of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2). Along with the rapid development of the single-cell RNA-sequencing (scRNA-seq) technology, the heterogeneity within the tumor microenvironment (TME) could be studied at a higher resolution level, facilitating an exploration of the mechanisms leading to poor prognosis during tumor progression. In previous studies, hypoxia was considered as an intrinsic characteristic of TME in solid tumors, which would activate downstream signaling pathways associated with angiogenesis and metastasis. Moreover, hypoxia-related genes (HRGs) based risk score models demonstrated nice performance in predicting the prognosis of TNBC patients. However, it is essential to further investigate the heterogeneity within hypoxic TME, such as intercellular communications. In the present study, utilizing single-sample Gene Set Enrichment Analysis (ssGSEA) and cell-cell communication analysis on the scRNA-seq data retrieved from Gene Expression Omnibus (GEO) database with accession number GSM4476488, we identified four tumor subpopulations with diverse functions, particularly a hypoxia-related one. Furthermore, results of cell-cell communication analysis revealed the dominant role of the hypoxic tumor subpopulation in angiogenesis- and metastasis-related signaling pathways as a signal sender. Consequently, regard the TNBC cohorts acquired from The Cancer Genome Atlas (TCGA) and GEO as train set and test set respectively, we constructed a risk score model with reliable capacity for the prediction of overall survival (OS), where ARTN and L1CAM were identified as risk factors promoting angiogenesis and metastasis of tumors. The expression of ARTN and L1CAM were further analyzed through tumor immune estimation resource (TIMER) platform. In conclusion, these two marker genes of the hypoxic tumor subpopulation played vital roles in tumor development, indicating poor prognosis in TNBC patients.
Citation: Yi Shi, Xiaoqian Huang, Zhaolan Du, Jianjun Tan. Analysis of single-cell RNA-sequencing data identifies a hypoxic tumor subpopulation associated with poor prognosis in triple-negative breast cancer[J]. Mathematical Biosciences and Engineering, 2022, 19(6): 5793-5812. doi: 10.3934/mbe.2022271
Triple-negative breast cancer (TNBC) is an aggressive subtype of mammary carcinoma characterized by low expression levels of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2). Along with the rapid development of the single-cell RNA-sequencing (scRNA-seq) technology, the heterogeneity within the tumor microenvironment (TME) could be studied at a higher resolution level, facilitating an exploration of the mechanisms leading to poor prognosis during tumor progression. In previous studies, hypoxia was considered as an intrinsic characteristic of TME in solid tumors, which would activate downstream signaling pathways associated with angiogenesis and metastasis. Moreover, hypoxia-related genes (HRGs) based risk score models demonstrated nice performance in predicting the prognosis of TNBC patients. However, it is essential to further investigate the heterogeneity within hypoxic TME, such as intercellular communications. In the present study, utilizing single-sample Gene Set Enrichment Analysis (ssGSEA) and cell-cell communication analysis on the scRNA-seq data retrieved from Gene Expression Omnibus (GEO) database with accession number GSM4476488, we identified four tumor subpopulations with diverse functions, particularly a hypoxia-related one. Furthermore, results of cell-cell communication analysis revealed the dominant role of the hypoxic tumor subpopulation in angiogenesis- and metastasis-related signaling pathways as a signal sender. Consequently, regard the TNBC cohorts acquired from The Cancer Genome Atlas (TCGA) and GEO as train set and test set respectively, we constructed a risk score model with reliable capacity for the prediction of overall survival (OS), where ARTN and L1CAM were identified as risk factors promoting angiogenesis and metastasis of tumors. The expression of ARTN and L1CAM were further analyzed through tumor immune estimation resource (TIMER) platform. In conclusion, these two marker genes of the hypoxic tumor subpopulation played vital roles in tumor development, indicating poor prognosis in TNBC patients.

| [1] |
F. Bray, J. Ferlay, I. Soerjomataram, R. L. Siegel, L. A. Torre, A. Jemal, Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries, CA Cancer J. Clin., 68 (2018), 394–424. https://doi.org/10.3322/caac.21492 doi: 10.3322/caac.21492

|
| [2] |
R. Dent, M. Trudeau, K. I. Pritchard, W. M. Hanna, H. K. Kahn, C. A. Sawka, et al., Triple-negative breast cancer: clinical features and patterns of recurrence, Clin. Cancer Res., 13 (2007), 4429–4434. https://doi.org/10.1158/1078-0432.CCR-06-3045 doi: 10.1158/1078-0432.CCR-06-3045
|
| [3] |
A. Marra, G. Viale, G. Curigliano, Recent advances in triple negative breast cancer: the immunotherapy era, BMC Medicine, 17 (2019), 90. https://doi.org/10.1186/s12916-019-1326-5 doi: 10.1186/s12916-019-1326-5
|
| [4] |
G. Zarrilli, G. Businello, M. V. Dieci, S. Paccagnella, V. Carraro, R. Cappellesso, et al., The tumor microenvironment of primitive and metastatic breast cancer: implications for novel therapeutic strategies, Int. J. Mol. Sci., 21 (2020), 8102. https://doi.org/10.3390/ijms21218102 doi: 10.3390/ijms21218102
|
| [5] |
S. D. Soysal, A. Tzankov, S. E. Muenst, Role of the tumor microenvironment in breast cancer, Pathobiology, 82 (2015), 142–152. https://doi.org/10.1159/000430499 doi: 10.1159/000430499
|
| [6] |
P. V. Loo, T. Voet, Single cell analysis of cancer genomes, Curr. Opin. Genet. Dev., 24 (2014), 82–91. https://doi.org/10.1016/j.gde.2013.12.004 doi: 10.1016/j.gde.2013.12.004
|
| [7] |
A. A. Pollen, T. J. Nowakowski, J. Shuga, X. Wang, A. A. Leyrat, J. H. Lui, et al., Low-coverage single-cell mRNA sequencing reveals cellular heterogeneity and activated signaling pathways in developing cerebral cortex, Nat. Biotechnol., 32 (2014), 1053–1058. https://doi.org/10.1038/nbt.2967 doi: 10.1038/nbt.2967
|
| [8] |
B. Treutlein, D. G. Brownfield, A. R. Wu, N. F. Neff, G. L. Mantalas, F. H. Espinoza, et al., Reconstructing lineage hierarchies of the distal lung epithelium using single-cell RNA-seq, Nature, 509 (2014), 371–375. https://doi.org/10.1038/nature13173 doi: 10.1038/nature13173
|
| [9] |
Q. H. Nguyen, N. Pervolarakis, K. Blake, D. Ma, R. T. Davis, N. James, et al., Profiling human breast epithelial cells using single cell RNA sequencing identifies cell diversity, Nat. Commun., 9 (2018), 2028. https://doi.org/10.1038/s41467-018-04334-1 doi: 10.1038/s41467-018-04334-1
|
| [10] |
M. Bartoschek, N. Oskolkov, M. Bocci, J. Lövrot, C. Larsson, M. Sommarin, et al., Spatially and functionally distinct subclasses of breast cancer-associated fibroblasts revealed by single cell RNA sequencing, Nat. Commun., 9 (2018), 5150. https://doi.org/10.1038/s41467-018-07582-3 doi: 10.1038/s41467-018-07582-3
|
| [11] |
D. Hanahan, R. A. Weinberg, Hallmarks of cancer: the next generation, Cell, 144 (2011), 646–674. https://doi.org/10.1016/j.cell.2011.02.013 doi: 10.1016/j.cell.2011.02.013
|
| [12] |
B. Muz, P. de la Puente, F. Azab, A. K. Azab, The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy, Hypoxia (Auckl), 3 (2015), 83–92. https://doi.org/10.2147/HP.S93413 doi: 10.2147/HP.S93413
|
| [13] |
X. Jing, F. Yang, C. Shao, K. Wei, M. Xie, H. Shen, et al., Role of hypoxia in cancer therapy by regulating the tumor microenvironment, Mol. Cancer, 18 (2019), 157. https://doi.org/10.1186/s12943-019-1089-9 doi: 10.1186/s12943-019-1089-9
|
| [14] |
X. Sun, H. Luo, C. Han, Y. Zhang, C. Yan, Identification of a hypoxia-related molecular classification and hypoxic tumor microenvironment signature for predicting the prognosis of patients with triple-negative breast cancer, Front. Oncol., 11 (2021), 700062. https://doi.org/10.3389/fonc.2021.700062 doi: 10.3389/fonc.2021.700062
|
| [15] |
X. Yang, X. Weng, Y. Yang, M. Zhang, Y. Xiu, W. Peng, et al., A combined hypoxia and immune gene signature for predicting survival and risk stratification in triple-negative breast cancer, Aging (Albany NY), 13 (2021), 19486–19509. https://doi.org/10.18632/aging.203360 doi: 10.18632/aging.203360
|
| [16] |
R. Gao, S. Bai, Y. C. Henderson, Y. Lin, A. Schalck, Y. Yan, et al., Delineating copy number and clonal substructure in human tumors from single-cell transcriptomes, Nat. Biotechnol., 39 (2021), 599–608. https://doi.org/10.1038/s41587-020-00795-2 doi: 10.1038/s41587-020-00795-2
|
| [17] |
M. J. Goldman, B. Craft, M. Hastie, K. Repečka, F. McDade, A. Kamath, et al., Visualizing and interpreting cancer genomics data via the Xena platform, Nat. Biotechnol., 38 (2020), 675–678. https://doi.org/10.1038/s41587-020-0546-8 doi: 10.1038/s41587-020-0546-8
|
| [18] |
D. M. Gendoo, N. Ratanasirigulchai, M. S. Schroder, L. Paré, J. S. Parker, A. Prat, et al., Genefu: an R/Bioconductor package for computation of gene expression-based signatures in breast cancer, Bioinformatics, 32 (2016), 1097–1099. https://doi.org/10.1093/bioinformatics/btv693 doi: 10.1093/bioinformatics/btv693
|
| [19] |
P. Jezequel, D. Loussouarn, C. Guerin-Charbonnel, L. Campion, A. Vanier, W. Gouraud, et al., Gene-expression molecular subtyping of triple-negative breast cancer tumours: importance of immune response, Breast Cancer Res., 17 (2015), 43. https://doi.org/10.1186/s13058-015-0550-y doi: 10.1186/s13058-015-0550-y
|
| [20] |
Y. Hao, S. Hao, E. Andersen-Nissen, W. M. Mauck III, S. Zheng, A. Butler, et al., Integrated analysis of multimodal single-cell data, Cell, 184 (2021), 3573–3587. https://doi.org/10.1016/j.cell.2021.04.048 doi: 10.1016/j.cell.2021.04.048
|
| [21] |
D. Aran, A. P. Looney, L. Liu, E. Wu, V. Fong, A. Hsu, et al., Reference-based analysis of lung single-cell sequencing reveals a transitional profibrotic macrophage, Nat. Immunol., 20 (2019), 163–172. https://doi.org/10.1038/s41590-018-0276-y doi: 10.1038/s41590-018-0276-y
|
| [22] |
A. Liberzon, C. Birger, H. Thorvaldsdottir, M. Ghandi, J. P. Mesirov, P. Tamayo, The Molecular Signatures Database (MSigDB) hallmark gene set collection, Cell Syst., 1 (2015), 417–425. https://doi.org/10.1016/j.cels.2015.12.004 doi: 10.1016/j.cels.2015.12.004
|
| [23] |
S. Jin, C. F. Guerrero-Juarez, L. Zhang, I. Chang, R. Ramos, C. H. Kuan, et al., Inference and analysis of cell-cell communication using CellChat, Nat. Commun., 12 (2021), 1088. https://doi.org/10.1038/s41467-021-21246-9 doi: 10.1038/s41467-021-21246-9
|
| [24] |
J. F. Prud'homme, F. Fridlansky, M. Le Cunff, M. Atger, C. Mercier-Bodart, M. F. Pichon, et al., Cloning of a gene expressed in human breast cancer and regulated by estrogen in MCF-7 cells, DNA, 4 (1985), 11–21. https://doi.org/10.1089/dna.1985.4.11 doi: 10.1089/dna.1985.4.11
|
| [25] |
M. C. Rio, J. P. Bellocq, J. Y. Daniel, C. Tomasetto, R. Lathe, M. P. Chenard, et al., Breast cancer-associated pS2 protein: synthesis and secretion by normal stomach mucosa, Science, 241 (1988), 705–708. https://doi.org/10.1126/science.3041593 doi: 10.1126/science.3041593
|
| [26] |
T. Vogl, A. Stratis, V. Wixler, T. Völler, S. Thurainayagam, S. K. Jorch, et al., Autoinhibitory regulation of S100A8/S100A9 alarmin activity locally restricts sterile inflammation, J. Clin. Invest., 128 (2018), 1852–1866. https://doi.org/10.1172/JCI89867 doi: 10.1172/JCI89867
|
| [27] |
Q. Fang, S. Yao, G. Luo, X. Zhang, Identification of differentially expressed genes in human breast cancer cells induced by 4-hydroxyltamoxifen and elucidation of their pathophysiological relevance and mechanisms, Oncotarget, 9 (2018), 2475–2501. https://doi.org/10.18632/oncotarget.23504 doi: 10.18632/oncotarget.23504
|
| [28] |
N. O'Brien, T. M. Maguire, N. O'Donovan, N. Lynch, A. D. Hill, E. McDermott, et al., Mammaglobin a: a promising marker for breast cancer, Clin. Chem., 48 (2002), 1362–1364. https://doi.org/10.1093/clinchem/48.8.1362 doi: 10.1093/clinchem/48.8.1362
|
| [29] |
M. Zafrakas, B. Petschke, A. Donner, F. Fritzsche, G. Kristiansen, R. Knüchel, et al., Expression analysis of mammaglobin A (SCGB2A2) and lipophilin B (SCGB1D2) in more than 300 human tumors and matching normal tissues reveals their co-expression in gynecologic malignancies, BMC Cancer, 6 (2006), 88. https://doi.org/10.1186/1471-2407-6-88 doi: 10.1186/1471-2407-6-88
|
| [30] |
D. Carter, J. F. Douglass, C. D. Cornellison, M. W. Retter, J. C. Johnson, A. A. Bennington, et al., Purification and characterization of the mammaglobin/lipophilin B complex, a promising diagnostic marker for breast cancer, Biochemistry, 41 (2002), 6714–6722. https://doi.org/10.1021/bi0159884 doi: 10.1021/bi0159884
|
| [31] |
T. L. Colpitts, P. Billing-Medel, P. Friedman, E. N. Granados, M. Hayden, S. Hodges, et al., Mammaglobin is found in breast tissue as a complex with BU101, Biochemistry, 40 (2001), 11048–11059. https://doi.org/10.1021/bi010284f doi: 10.1021/bi010284f
|
| [32] |
S. Robson, S. Pelengaris, M. Khan, c-Myc and downstream targets in the pathogenesis and treatment of cancer, Recent Pat. Anticancer Drug Discov., 1 (2006), 305–326. https://doi.org/10.2174/157489206778776934 doi: 10.2174/157489206778776934
|
| [33] |
P. C. Fernandez, S. R. Frank, L. Wang, M. Schroeder, S. Liu, J. Greene, et al., Genomic targets of the human c-Myc protein, Genes Dev., 17 (2003), 1115–1129. https://doi.org/10.1101/gad.1067003 doi: 10.1101/gad.1067003
|
| [34] |
J. H. Patel, A. P. Loboda, M. K. Showe, L. C. Showe, S. B. McMahon, Analysis of genomic targets reveals complex functions of MYC, Nat. Rev. Cancer, 4 (2004), 562–568. https://doi.org/10.1038/nrc1393 doi: 10.1038/nrc1393
|
| [35] |
C. Attwooll, E. L. Denchi, K. Helin, The E2F family: specific functions and overlapping interests, EMBO J., 23 (2004), 4709–4716. https://doi.org/10.1038/sj.emboj.7600481 doi: 10.1038/sj.emboj.7600481
|
| [36] |
T. Yu, L. Liang, X. Zhao, Y. Yin, Structural and biochemical studies of the extracellular domain of Myelin protein zero-like protein 1, Biochem. Biophys. Res. Commun., 506 (2018), 883–890. https://doi.org/10.1016/j.bbrc.2018.10.161 doi: 10.1016/j.bbrc.2018.10.161
|
| [37] |
K. M. McCarthy, I. B. Skare, M. C. Stankewich, M. Furuse, S. Tsukita, R. A. Rogers, et al., Occludin is a functional component of the tight junction, J. Cell Sci., 109 (1996), 2287–2298. https://doi.org/10.1242/jcs.109.9.2287 doi: 10.1242/jcs.109.9.2287
|
| [38] |
J. Sakata, T. Shimokubo, K. Kitamura, S. Nakamura, K. Kangawa, H. Matsuo, et al., Molecular cloning and biological activities of rat adrenomedullin, a hypotensive peptide, Biochem. Biophys. Res. Commun., 195 (1993), 921–927. https://doi.org/10.1006/bbrc.1993.2132 doi: 10.1006/bbrc.1993.2132
|
| [39] |
K. Miyashita, H. Itoh, N. Sawada, Y. Fukunaga, M. Sone, K. Yamahara, et al., Adrenomedullin promotes proliferation and migration of cultured endothelial cells, Hypertens. Res., 26 Suppl (2003), S93–98. https://doi.org/10.1291/hypres.26.S93 doi: 10.1291/hypres.26.S93
|
| [40] |
N. Ferrara, H. P. Gerber, J. LeCouter, The biology of VEGF and its receptors, Nat. Med., 9 (2003), 669–676. https://doi.org/10.1038/nm0603-669 doi: 10.1038/nm0603-669
|
| [41] |
M. I. Lin, W. C. Sessa, Vascular endothelial growth factor signaling to endothelial nitric oxide synthase: more than a FLeeTing moment, Circ. Res., 99 (2006), 666–668. https://doi.org/10.1161/01.RES.0000245430.24075.a4 doi: 10.1161/01.RES.0000245430.24075.a4
|
| [42] |
S. Sugo, N. Minamino, K. Kangawa, K. Miyamoto, K. Kitamura, J. Sakata, et al., Endothelial cells actively synthesize and secrete adrenomedullin, Biochem. Biophys. Res. Commun., 201 (1994), 1160–1166. https://doi.org/10.1006/bbrc.1994.1827 doi: 10.1006/bbrc.1994.1827
|
| [43] |
M. J. Miller, A. Martinez, E. J. Unsworth, C. J. Thiele, T. W. Moody, T. Elsasser, et al., Adrenomedullin expression in human tumor cell lines. Its potential role as an autocrine growth factor, J. Biol. Chem., 271 (1996), 23345–23351. https://doi.org/10.1074/jbc.271.38.23345 doi: 10.1074/jbc.271.38.23345
|
| [44] | K. Dawas, M. Loizidou, A. Shankar, H. Ali, I. Taylor, Angiogenesis in cancer: the role of endothelin-1, Ann. R. Coll. Surg. Engl., 81 (1999), 306–310. |
| [45] |
N. Zhu, L. Gu, J. Jia, X. Wang, L. Wang, M. Yang, W. Yuan, Endothelin-1 triggers human peritoneal mesothelial cells' proliferation via ERK1/2-Ets-1 signaling pathway and contributes to endothelial cell angiogenesis, J. Cell. Biochem., 120 (2019), 3539–3546. https://doi.org/10.1002/jcb.27631 doi: 10.1002/jcb.27631
|
| [46] |
Y. Katanasaka, Y. Kodera, Y. Kitamura, T. Morimoto, T. Tamura, F. Koizumi, Epidermal growth factor receptor variant type III markedly accelerates angiogenesis and tumor growth via inducing c-myc mediated angiopoietin-like 4 expression in malignant glioma, Mol. Cancer, 12 (2013), 31. https://doi.org/10.1186/1476-4598-12-31 doi: 10.1186/1476-4598-12-31
|
| [47] |
R. Kolb, P. Kluz, Z. W. Tan, N. Borcherding, N. Bormann, A. Vishwakarma, et al., Obesity-associated inflammation promotes angiogenesis and breast cancer via angiopoietin-like 4, Oncogene, 38 (2019), 2351–2363. https://doi.org/10.1038/s41388-018-0592-6 doi: 10.1038/s41388-018-0592-6
|
| [48] |
M. S. Airaksinen, M. Saarma, The GDNF family: signalling, biological functions and therapeutic value, Nat. Rev. Neurosci., 3 (2002), 383–394. https://doi.org/10.1038/nrn812 doi: 10.1038/nrn812
|
| [49] |
J. Kang, P. X. Qian, V. Pandey, J. K. Perry, L. D. Miller, E. T. Liu, et al., Artemin is estrogen regulated and mediates antiestrogen resistance in mammary carcinoma, Oncogene, 29 (2010), 3228–3240. https://doi.org/10.1038/onc.2010.71 doi: 10.1038/onc.2010.71
|
| [50] |
J. Kang, J. K. Perry, V. Pandey, G. C. Fielder, B. Mei, P. X. Qian, et al., Artemin is oncogenic for human mammary carcinoma cells, Oncogene, 28 (2009), 2034–2045. https://doi.org/10.1038/onc.2009.66 doi: 10.1038/onc.2009.66
|
| [51] |
A. Banerjee, Z. S. Wu, P. Qian, J. Kang, V. Pandey, D. X. Liu, et al., ARTEMIN synergizes with TWIST1 to promote metastasis and poor survival outcome in patients with ER negative mammary carcinoma, Breast Cancer Res., 13 (2011), R112. https://doi.org/10.1186/bcr3054 doi: 10.1186/bcr3054
|
| [52] |
A. Banerjee, P. Qian, Z. S. Wu, X. Ren, M. Steiner, N. M. Bougen, et al., Artemin stimulates radio- and chemo-resistance by promoting TWIST1-BCL-2-dependent cancer stem cell-like behavior in mammary carcinoma cells, J. Biol. Chem., 287 (2012), 42502–42515. https://doi.org/10.1074/jbc.M112.365163 doi: 10.1074/jbc.M112.365163
|
| [53] |
H. Zhang, C. C. Wong, H. Wei, D. M. Gilkes, P. Korangath, P. Chaturvedi, et al., HIF-1-dependent expression of angiopoietin-like 4 and L1CAM mediates vascular metastasis of hypoxic breast cancer cells to the lungs, Oncogene, 31 (2012), 1757–1770. https://doi.org/10.1038/onc.2011.365 doi: 10.1038/onc.2011.365
|
| [54] |
A. Banerjee, Z. S. Wu, P. X. Qian, J. Kang, D. X. Liu, T. Zhu, et al., ARTEMIN promotes de novo angiogenesis in ER negative mammary carcinoma through activation of TWIST1-VEGF-A signaling, PLoS One, 7 (2012), e50098. https://doi.org/10.1371/journal.pone.0050098 doi: 10.1371/journal.pone.0050098
|
| [55] |
A. Friedli, E. Fischer, Novak-Hofer I, S. Cohrs, K. Ballmer-Hofer, P. A. Schubiger, et al., The soluble form of the cancer-associated L1 cell adhesion molecule is a pro-angiogenic factor, Int. J. Biochem. Cell Biol., 41 (2009), 1572–1580. https://doi.org/10.1016/j.biocel.2009.01.006 doi: 10.1016/j.biocel.2009.01.006
|
| [56] |
H. Hall, J. A. Hubbell, Matrix-bound sixth Ig-like domain of cell adhesion molecule L1 acts as an angiogenic factor by ligating alphavbeta3-integrin and activating VEGF-R2, Microvasc. Res., 68 (2004), 169–178. https://doi.org/10.1016/j.mvr.2004.07.001 doi: 10.1016/j.mvr.2004.07.001
|
| [57] |
H. Hall, V. Djonov, M. Ehrbar, M. Hoechli, J. A. Hubbell, Heterophilic interactions between cell adhesion molecule L1 and alphavbeta3-integrin induce HUVEC process extension in vitro and angiogenesis in vivo, Angiogenesis, 7 (2004), 213–223. https://doi.org/10.1007/s10456-004-1328-5 doi: 10.1007/s10456-004-1328-5
|
| [58] |
M. Zhang, W. Zhang, Z. Wu, S. Liu, L. Sun, Y. Zhong, et al., Artemin is hypoxia responsive and promotes oncogenicity and increased tumor initiating capacity in hepatocellular carcinoma, Oncotarget, 7 (2016), 3267–3282. https://doi.org/10.18632/oncotarget.6572 doi: 10.18632/oncotarget.6572
|









Figures(10) / Tables(1)

Yi Shi, Xiaoqian Huang, Zhaolan Du, Jianjun Tan. Analysis of single-cell RNA-sequencing data identifies a hypoxic tumor subpopulation associated with poor prognosis in triple-negative breast cancer[J]. Mathematical Biosciences and Engineering, 2022, 19(6): 5793-5812. doi: 10.3934/mbe.2022271







 DownLoad:
DownLoad: