Citation: Kourosh Cheraghipour, Abdolrazagh Marzban, Behrouz Ezatpour, Sayyad Khanizadeh, Javad Koshki. Antiparasitic properties of curcumin: A review[J]. AIMS Agriculture and Food, 2018, 3(4): 561-578. doi: 10.3934/agrfood.2018.4.561

| [1] | Croft SL, Sundar S, Fairlamb AH (2006) Drug resistance in leishmaniasis. Clin Microbiol Rev 19: 111–126. |
| [2] | Hayat F, Azam A, Shin D (2016) Recent progress on the discovery of antiamoebic agents. Bioorg Med Chem Lett 26: 5149–5159. |
| [3] | Oryan A, Akbari M (2016) Worldwide risk factors in leishmaniasis. Asian Pac J Trop Med 9: 925–932. |
| [4] | Aggarwal BB, Sundaram C, Malani N, et al. (2007) Curcumin: The Indian solid gold, In: The molecular targets and therapeutic uses of curcumin in health and disease, 1–75. |
| [5] | Agarwal A, Saxena PN (2018) Curcumin, a polyphenol from the Curcuma longa, prevents mercuric chloride-induced liver damage through reversal of oxidative stress and biochemical changes. AACR. |
| [6] | Aggarwal BB, Kumar A, Bharti AC (2003) Anticancer potential of curcumin: preclinical and clinical studies. Anticancer Res 23: 363–398. |
| [7] | Galasso V, Kovac B, Modelli A, et al. (2008) Spectroscopic and theoretical study of the electronic structure of curcumin and related fragment molecules. J Phys Chem A 112: 2331–2338. |
| [8] | Sun J, Zhao Y, Hu J (2013) Curcumin inhibits imiquimod-induced psoriasis-like inflammation by inhibiting IL-1beta and IL-6 production in mice. PloS one 8: e67078. |
| [9] | Aggarwal BB, Sung B (2009) Pharmacological basis for the role of curcumin in chronic diseases: An age-old spice with modern targets. Trends Pharmacol Sci 30: 85–94. |
| [10] | Strimpakos AS, Sharma RA (2008) Curcumin: Preventive and therapeutic properties in laboratory studies and clinical trials. Antioxid Redox Signal 10: 511–546. |
| [11] | Chen C, Johnston TD, Jeon H, et al. (2009) An in vitro study of liposomal curcumin: Stability, toxicity and biological activity in human lymphocytes and Epstein-Barr virus-transformed human B-cells. Int J Pharm 366: 133–139. |
| [12] | Asadpour M, Namazi F, Razavi SM, et al. (2018) Comparative efficacy of curcumin and paromomycin against Cryptosporidium parvum infection in a BALB/c model. Vet Parasitol 250: 7–14. |
| [13] | Feng J, Zhang L, Huang F, et al. (2018) Ready for malaria elimination: Zero indigenous case reported in the People's Republic of China. Malar J 17: 315. |
| [14] | Martinez A, Janitschke K (1985) Acanthamoeba, an opportunistic microorganism: A review. Infection 13: 251–256. |
| [15] | Baig AM (2015) Pathogenesis of amoebic encephalitis: Are the amoebae being credited to an "inside job" done by the host immune response? Acta Trop 148: 72–76. |
| [16] | Petri Jr W, Haque R, Lyerly D, et al. (2000) Estimating the impact of amebiasis on health. Parasitol Today 16: 320–321. |
| [17] | McClelland RS, Sangaré L, Hassan WM, et al. (2007) Infection with Trichomonas vaginalis increases the risk of HIV-1 acquisition. J Infect Dis 195: 698–702. |
| [18] | Minetti C, Chalmers RM, Beeching NJ, et al. (2016) Giardiasis. BMJ 40: 241–251. |
| [19] | Guy RA, Xiao C, Horgen PA (2004) Real-time PCR assay for detection and genotype differentiation of Giardia lamblia in stool specimens. J Clin Microbiol 42: 3317–3320. |
| [20] | Tenter AM, Heckeroth AR, Weiss LM (2000) Toxoplasma gondii: From animals to humans. Int J Parasitol 30: 1217–1258. |
| [21] | Cook AJC, Holliman R, Gilbert RE, et al. (2000) Sources of Toxoplasma infection in pregnant women: European multicentre case-control study Commentary: Congenital Toxoplasmosis—further thought for food. BMJ 321: 142–147. |
| [22] | Qian W, Wang H, Shan D, et al. (2015) Activity of several kinds of drugs against Neospora caninum. Parasitol Int 64: 597–602. |
| [23] | Dubey J, Lindsay D (1996) A review of Neospora caninum and neosporosis. Vet Parasitol 67: 1–59. |
| [24] | Checkley W, Epstein LD, Gilman RH, et al. (1998) Effects of Cryptosporidium parvum infection in Peruvian children: Growth faltering and subsequent catch-up growth. Am J Epidemiol 148: 497–506. |
| [25] | Kramer MH, Herwaldt BL, Craun GF, et al. (1996) Surveillance for waterborne-disease outbreaks—United States, 1993–1994. MMWR CDC Surveill Summ 12: 1–33. |
| [26] | Lee SH, Lillehoj HS, Jang SI, et al. (2011) Effects of dietary supplementation with phytonutrients on vaccine-stimulated immunity against infection with Eimeria tenella. Vet Parasitol 181: 97–105. |
| [27] | Giannenas I, Florou-Paneri P, Papazahariadou M, et al. (2003) Effect of dietary supplementation with oregano essential oil on performance of broilers after experimental infection with Eimeria tenella. Arch Anim Nutr 57: 99–106. |
| [28] | Novaes RD, Sartini MVP, Rodrigues JPF, et al. (2016) Curcumin enhances the anti-Trypanosoma cruzi activity of benznidazole-based chemotherapy in acute experimental Chagas disease. Antimicrob Agents Chemother 60: 3355–3364. |
| [29] | Sueth-Santiago V, de BB Moraes J, Alves ESS, et al. (2016) The effectiveness of natural diarylheptanoids against Trypanosoma cruzi: Cytotoxicity, ultrastructural alterations and molecular modeling studies. PLoS One 11: e0162926. |
| [30] | Baker J (1995) The subspecific taxonomy of Trypanosoma brucei. Parasite 2: 3–12. |
| [31] | Deborggraeve S, Koffi M, Jamonneau V, et al. (2008) Molecular analysis of archived blood slides reveals an atypical human Trypanosoma infection. Diagn Micr Infect Dis 61: 428–433. |
| [32] | Hussein A, Rashed S, El Hayawan I, et al. (2017) Evaluation of the Anti-schistosomal Effects of Turmeric (Curcuma longa) Versus Praziquantel in Schistosoma mansoni Infected Mice. Iran J Parasitol 12: 587–596. |
| [33] | Oluwole AS, Adeniran AA, Mogaji HO, et al. (2018) Prevalence, intensity and spatial co-distribution of schistosomiasis and soil transmitted helminths infections in Ogun state, Nigeria. Parasitol Open 4. |
| [34] | Sarkari B, Khabisi SA (2017) Immunodiagnosis of Human Fascioliasis: An Update of Concepts and Performances of the Serological Assays. J Clin Diagn Res 11: OE05–OE10. |
| [35] | Bauri R, Tigga MN, Kullu SS (2015) A review on use of medicinal plants to control parasites. Indian J Nat Prod Resour 6: 268–277. |
| [36] | Wink M (2012) Medicinal plants: A source of anti-parasitic secondary metabolites. Molecules 17: 12771–12791. |
| [37] | Hatcher H, Planalp R, Cho J, et al. (2008) Curcumin: From ancient medicine to current clinical trials. Cell Mol Life Sci 65: 1631–1652. |
| [38] | Mohammad BA (2011) Antileishmanial effects of traditional herbal extracts against cutaneous leishmaniosis in vivo. Adv Environ Biol 5: 3188–3195. |
| [39] | Hosseini A, Jaffary F, Asghari GR, et al. (2012) In Vitro Effects of Turmeric and Licorice Total Extracts on L. Major Promastigotes. J Isfahan Med Sch 29: 1–11. |
| [40] | Khanra S, Kumar YP, Dash J, et al. (2018) In vitro screening of known drugs identified by scaffold hopping techniques shows promising leishmanicidal activity for suramin and netilmicin. BMC Res Notes 11: 319. |
| [41] | Amaral ACF, Gomes LA, Silva JRdA, et al. (2014) Liposomal formulation of turmerone-rich hexane fractions from Curcuma longa enhances their antileishmanial activity. BioMed Res Int 2014: 694934. |
| [42] | Fouladvand M, Barazesh A, Tahmasebi R (2013) Evaluation of in vitro antileishmanial activity of curcumin and its derivatives "gallium curcumin, indium curcumin and diacethyle curcumin". Eur Rev Med Pharmacol Sci 17: 3306–3308. |
| [43] | Tiwari B, Pahuja R, Kumar P, et al. (2017) Nanotized curcumin and miltefosine, a potential combination for treatment of experimental visceral leishmaniasis. Antimicrob Agents Chemother 61: e01169-16. |
| [44] | Busari ZA, Dauda KA, Morenikeji OA, et al. (2017) Antiplasmodial Activity and Toxicological Assessment of Curcumin PLGA-Encapsulated Nanoparticles. Front Pharmacol 8: 622. |
| [45] | Donipati P, Hara Sreeramulu S (2015) In vitro anti-malarial activity of Rhizome extracts of Curcuma species. Int J Pharm Bio Sci 5: 488–494. |
| [46] | Vathsala PG, Dende C, Nagaraj VA, et al. (2012) Curcumin-arteether combination therapy of Plasmodium berghei-infected mice prevents recrudescence through immunomodulation. PLoS One 7: e29442. |
| [47] | Singh DB, Dwivedi S (2016) Structural insight into binding mode of inhibitor with SAHH of Plasmodium and human: interaction of curcumin with anti-malarial drug targets. J Chem Biol 9: 107–120. |
| [48] | Dende C, Meena J, Nagarajan P, et al. (2015) Simultaneously targeting inflammatory response and parasite sequestration in brain to treat Experimental Cerebral Malaria. Sci Rep 5: 12671. |
| [49] | Alam S, Panda JJ, Mukherjee TK, et al. (2016) Short peptide based nanotubes capable of effective curcumin delivery for treating drug resistant malaria. J Nanobiotechnol 14: 26. |
| [50] | Akhtar F, Rizvi MMA, Kar SK (2012) Oral delivery of curcumin bound to chitosan nanoparticles cured Plasmodium yoelii infected mice. Biotechnol Adv 30: 310–320. |
| [51] | Ghosh A, Banerjee T, Bhandary S, et al. (2014) Formulation of nanotized curcumin and demonstration of its antimalarial efficacy. Int J Nanomedicine 9: 5373–5387. |
| [52] | Kunwittaya S, Treeratanapiboon L, Srisarin A, et al. (2014) In vitro study of parasite elimination and endothelial protection by curcumin: Adjunctive therapy for cerebral malaria. EXCLI J 13: 287–299. |
| [53] | Dohutia C, Chetia D, Gogoi K, et al. (2017) Design, in silico and in vitro evaluation of curcumin analogues against Plasmodium falciparum. Exp Parasitol 175: 51–58. |
| [54] | El-Sayed NM, Ismail KA, Ahmed SAEG, et al. (2012) In vitro amoebicidal activity of ethanol extracts of Arachis hypogaea L., Curcuma longa L. and Pancratium maritimum L. on Acanthamoeba castellanii cysts. Parasitol Res 110: 1985–1992. |
| [55] | Rangel‐Castañeda IA, Hernández‐Hernández JM, Pérez‐Rangel A, et al. (2018) Amoebicidal activity of curcumin on Entamoeba histolytica trophozoites. J Pharm Pharmacol 70: 426–433. |
| [56] | Wachter B, Syrowatka M, Obwaller A, et al. (2014) In vitro efficacy of curcumin on Trichomonas vaginalis. Wien Klin Wochenschr 126: 32–36. |
| [57] | Gutiérrez-Gutiérrez F, Palomo-Ligas L, Hernández-Hernández JM, et al. (2017) Curcumin alters the cytoskeleton and microtubule organization on trophozoites of Giardia lamblia. Acta Trop 172: 113–121. |
| [58] | Said D, Elsamad L, Gohar Y (2012) Validity of silver, chitosan, and curcumin nanoparticles as anti-Giardia agents. Parasitol Res 111: 545–554. |
| [59] | Goo YK, Yamagishi J, Ueno A, et al. (2015) Characterization of Toxoplasma gondii glyoxalase 1 and evaluation of inhibitory effects of curcumin on the enzyme and parasite cultures. Parasites Vectors 8: 654. |
| [60] | Kim DK, Lillehoj HS, Lee SH, et al. (2013) Dietary Curcuma longa enhances resistance against Eimeria maxima and Eimeria tenella infections in chickens. Poultry Sci 92: 2635–2643. |
| [61] | Khalafalla RE, Müller U, Shahiduzzaman M, et al. (2011) Effects of curcumin (diferuloylmethane) on Eimeria tenella sporozoites in vitro. Parasit Res 108: 879–886. |
| [62] | Burt S, Tersteeg-Zijderveld M, Jongerius-Gortemaker B, et al. (2013) In vitro inhibition of Eimeria tenella invasion of epithelial cells by phytochemicals. Vet Parasitol 191: 374–378. |
| [63] | Nagajyothi F, Zhao D, Weiss LM, et al. (2012) Curcumin treatment provides protection against Trypanosoma cruzi infection. Parasitol Res 110: 2491–2499. |
| [64] | Hernández M, Wicz S, Corral RS (2016) Cardioprotective actions of curcumin on the pathogenic NFAT/COX-2/prostaglandin E2 pathway induced during Trypanosoma cruzi infection. Phytomedicine 23: 1392–1400. |
| [65] | de Paula Aguiar D, Moscardini MBM, Morais ER, et al. (2016) Curcumin Generates Oxidative Stress and Induces Apoptosis in Adult Schistosoma mansoni Worms. PLoS One 11: e0167135. |
| [66] | Morais ER, Oliveira KC, Magalhães LG, et al. (2013) Effects of curcumin on the parasite Schistosoma mansoni: A transcriptomic approach. Mol Biochem Parasit 187: 91–97. |
| [67] | Ullah R, Rehman A, Zafeer MF, et al. (2017) Anthelmintic potential of thymoquinone and curcumin on Fasciola gigantica. PLoS One 12: e0171267. |
| [68] | Oyeyemi O, Adegbeyeni O, Oyeyemi I, et al. (2018) In vitro ovicidal activity of poly lactic acid curcumin-nisin co-entrapped nanoparticle against Fasciola spp. eggs and its reproductive toxicity. J Basic Clin Physiol Pharmacol 29: 73–79. |
| [69] | Desta A, Shiferaw S, Kassa A, et al. (2005) Module on Leishmaniasis for the Ethiopian Health Center Team. Debub University, Ethiopia. |
| [70] | Ameen M (2007) Cutaneous leishmaniasis: Therapeutic strategies and future directions. Expert Opin Pharmacother 8: 2689–2699. |
| [71] | Lehane AM, Kirk K (2010) Efflux of a range of antimalarial drugs and "chloroquine resistance reversers" from the digestive vacuole in malaria parasites with mutant PfCRT. Mol Microbiol 77: 1039–1051. |
| [72] | Padmanaban G, Nagaraj VA, Rangarajan PN (2012) Artemisinin-based combination with curcumin adds a new dimension to malaria therapy. Curr Sci 102: 704–711. |
| [73] | Cui L, Miao J, Cui L (2007) Cytotoxic effect of curcumin on malaria parasite Plasmodium falciparum: Inhibition of histone acetylation and generation of reactive oxygen species. Antimicrob Agents Chemother 51: 488–494. |
| [74] | Obeid WN, de Araújo R, Vieira LA, et al. (2003) Acanthamoeba bilateral keratiti—Case report. Rev Bras Oftalmol 66: 876–880. |
| [75] | Farthing MJ (2006) Treatment options for the eradication of intestinal protozoa. Nat Rev Gastroenterol Hepatol 3: 436-445. |
| [76] | Meidan I, Sellam G, Simaan S, et al. (2013) Topical curcumin for the prevention of oral mucositis in pediatric patients: Case series. Altern Ther Health Med 19: 21-24. |
| [77] | Granados CE, Reveiz L, Uribe LG, et al. (2012) Drugs for treating giardiasis. Cochrane database of systematic reviews 12: CD007787. |
| [78] | Gilbert R, Gras L, Wallon M, et al. (2001) Effect of prenatal treatment on mother to child transmission of Toxoplasma gondii: Retrospective cohort study of 554 mother-child pairs in Lyon, France. Int J Epidemiol 30: 1303–1308. |
| [79] | Mofenson LM, Brady MT, Danner SP, et al. (2009) Guidelines for the prevention and treatment of opportunistic infections among HIV-exposed and HIV-infected children: Recommendations from CDC, the National Institutes of Health, the HIV Medicine Association of the Infectious Diseases Society of America, the Pediatric Infectious Diseases Society, and the American Academy of Pediatrics. MMWR Recomm Rep 58: 1–198. |
| [80] | Stephen LE (1986) Trypanosomiasis: A veterinary perspective. Pergamon Press, Oxford. |
| [81] | Babokhov P, Sanyaolu AO, Oyibo WA, et al. (2013) A current analysis of chemotherapy strategies for the treatment of human African trypanosomiasis. Pathog Global Health 107: 242–252. |
| [82] | Kowalska A, Kowalski P, Torres MÁT (2011) Chagas disease–american trypanosomiasis. Pol Ann Med 18: 156–167. |
| [83] | Zanolla D, Perissutti B, Passerini N, et al. (2018) Milling and comilling Praziquantel at cryogenic and room temperatures: Assessment of the process-induced effects on drug properties. J Pharmaceut Biomed 153: 82–89. |
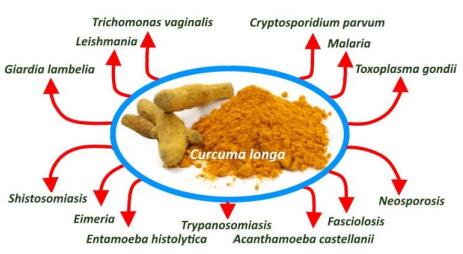
Figures(1) / Tables(2)

Kourosh Cheraghipour, Abdolrazagh Marzban, Behrouz Ezatpour, Sayyad Khanizadeh, Javad Koshki. Antiparasitic properties of curcumin: A review[J]. AIMS Agriculture and Food, 2018, 3(4): 561-578. doi: 10.3934/agrfood.2018.4.561







 DownLoad:
DownLoad: