Citation: Changyong Xu, Qiang Li, Tonghua Zhang, Sanling Yuan. Stability and Hopf bifurcation for a delayed diffusive competition model with saturation effect[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 8037-8051. doi: 10.3934/mbe.2020407

| [1] |
K. Gopalsamy, P. Weng, Global attractivity in a competition system with feedback controls, Comput. Math. Appl., 45 (2003), 665-676. doi: 10.1016/S0898-1221(03)00026-9

|
| [2] |
X. Tang, X. Zou, 3/2-type criteria for global attractivity of Lotka-Volterra competition system without instantaneous negative feedbacks, J. Differ. Equations, 186 (2002), 420-439. doi: 10.1016/S0022-0396(02)00011-6
|
| [3] |
J. Zhang, Z. Jin, J. Yan, G. Sun, Stability and Hopf bifurcation in a delayed competition system, Nonlinear Anal., 70 (2009), 658-670. doi: 10.1016/j.na.2008.01.002
|
| [4] |
P. Weng, Global attractivity in a periodic competition system with feedback controls, Acta. Math. Appl. Sin., 12 (1996), 11-21. doi: 10.1007/BF02009556
|
| [5] | Y. Li, Positive periodic solutions of discrete Lotka-Volterra competition systems with state dependent and distributed delays, Appl. Math. Comput., 190 (2007), 526-531. |
| [6] | J. Qiu, J. Cao, Exponential stability of a competitive Lotka-Volterra system with delays, Appl. Math. Comput., 201 (2008), 819-829. |
| [7] |
Y. Zhao, S. Yuan, J. Ma, Survival and stationary distribution analysis of a stochastic competitive model of three species in a polluted environment, Bull. Math. Biol., 77 (2015), 1285-1326. doi: 10.1007/s11538-015-0086-4
|
| [8] |
C. Xu, S. Yuan, Competition in the chemostat: a stochastic multi-species model and its asymptotic behavior, Math. Biosci., 280 (2016), 1-9. doi: 10.1016/j.mbs.2016.07.008
|
| [9] | Y. Xiao, Y. Zhou, S. Tang, Principles of biological mathematics, Xian Jiaotong University Press, Xian, 2012. |
| [10] |
Z. Jin, Z. Ma, Stability for a competitive Lotka-Volterra system with delays, Nonlinear Anal. Theory, Methods Appl., 51 (2002), 1131-1142. doi: 10.1016/S0362-546X(01)00881-1
|
| [11] |
S. Yuan, D. Wu, G. Lan, H. Wang, Noise-induced transitions in a nonsmooth producer-grazer model with stoichiometric constraints, B. Math. Biol., 82 (2020), 55. doi: 10.1007/s11538-020-00733-y
|
| [12] |
Z. Lu, Y. Takeuchi, Permanence and global attractivity for competitive Lotka-Volterra system with delay, Nonlinear Anal. Theory, Methods Appl., 22 (1994), 847-856. doi: 10.1016/0362-546X(94)90053-1
|
| [13] |
L. Zhou, Y. Tang, S. Hussein, Stability and Hopf bifurcation for a delay competition diffusion system, Chaos Solit. Fract., 14 (2002), 1201-1225. doi: 10.1016/S0960-0779(02)00068-1
|
| [14] |
Y. Song, M. Han, Y. Peng, Stability and Hopf bifurcations in a competitive Lotka-Volterra system with two delays, Chaos Solit. Fract., 22 (2004), 1139-1148. doi: 10.1016/j.chaos.2004.03.026
|
| [15] | X. Yu, S. Yuan, Asymptotic properties of a stochastic chemostat model with two distributed delays and nonlinear perturbation, Discrete Contin. Dyn. Syst. Ser. B, 25 (2020), 2373-2390. |
| [16] |
S. Zhao, S. Yuan, H. Wang, Threshold behavior in a stochastic algal growth model with stoichiometric constraints and seasonal variation, J. Differ. Equations, 268 (2020), 5113-5139. doi: 10.1016/j.jde.2019.11.004
|
| [17] | K. Gopalsamy, Stability and oscillations in delay differential equations of population dynamics, Springer, Netherlands, 1992. |
| [18] |
Z. Liu, R. Tan, Y. Chen, Modeling and analysis of a delayed competitive system with impulsive perturbations, Rocky Mount. J. Math., 38 (2008), 1505-1523. doi: 10.1216/RMJ-2008-38-5-1505
|
| [19] |
Z. Liu, J. Wu, R. Tan, Permanence and extinction of an impulsive delay competitive Lotka-Volterra model with periodic coefficients, IMA J. Appl. Math., 74 (2009), 559-573. doi: 10.1093/imamat/hxp007
|
| [20] |
G. Sun, Z. Jin, Y. Zhao, Q. Liu, L. Li, Spatial pattern in a predator prey system with both self-and cross-diffusion, Int. J. Mod. Phy. C, 20 (2009), 71-84. doi: 10.1142/S0129183109013467
|
| [21] | Q. Li, S. Yuan, Cross-Diffusion induced Turing instability for a competition model with saturation effect, Appl. Math. Compu., 347 (2019), 64-77. |
| [22] |
D. Jia, T. Zhang, S. Yuan, Pattern dynamics of a diffusive toxin producing phytoplanktonzooplankton model with three-dimensional patch, Int. J. Bifurcat. Chaos, 29 (2019), 1930011. doi: 10.1142/S0218127419300118
|
| [23] |
X. Yu, S. Yuan, T. Zhang, Asymptotic properties of stochastic nutrient-plankton food chain models with nutrient recycling, Nonlinear Anal. Hybri., 34 (2019), 209-225. doi: 10.1016/j.nahs.2019.06.005
|
| [24] |
J. Yang, T. Zhang, S. Yuan, Turing pattern induced by cross-diffusion in a predator-prey model with pack predation-herd behavior, Int. J. Bifurcat. Chaos, 30 (2020), 2050103. doi: 10.1142/S0218127420501035
|
| [25] |
C. Xu, S. Yuan, and T. Zhang, Global dynamics of a predator-prey model with defence mechanism for prey, Appl. Math. Lett., 62 (2016), 42-48. doi: 10.1016/j.aml.2016.06.013
|
| [26] |
G. Sun, C. Wang, Z. Wu, Pattern dynamics of a Gierer CMeinhardt model with spatial effects, Nonlinear Dynam., 88 (2017), 1385-1396. doi: 10.1007/s11071-016-3317-9
|
| [27] |
T. Zhang, Z. Hong, Delay-induced Turing instability in reaction-diffusion equations, Phys. Rev. E, 90 (2014), 052908. doi: 10.1103/PhysRevE.90.052908
|
| [28] |
J. Li, G. Sun, Z. Jin, Pattern formation of an epidemic model with time delay, Phys. A, 403 (2014), 100-109. doi: 10.1016/j.physa.2014.02.025
|
| [29] | X. Lian, H. Wang, W. Wang, Delay-driven pattern formation in a reaction-diffusion predator-prey model incorporating a prey refuge, J. Stat. Mech. Theory Exp., 2013 (2013), P04006. |
| [30] |
E. Beretta, Y. Kuang, Global analyses in some delayed ratio-dependent predator-prey systems, Nonlinear Anal., 32 (1998), 381-408 doi: 10.1016/S0362-546X(97)00491-4
|
| [31] |
S. Hsu, T. Hwang, Y. Kuang, Global analysis of the Michaelis-Menten type ratio-dependent predator-prey system, J. Math. Biol., 42 (2001), 489-506. doi: 10.1007/s002850100079
|
| [32] |
T. Faria, Normal forms and Hopf bifurcation for partial differential equations with delays, T. Am. Math. Soc., 352 (2000), 2217-2238. doi: 10.1090/S0002-9947-00-02280-7
|
| [33] |
T. Faria, Stability and bifurcation for a delayed predator-prey model and the effect of diffusion, J. Math. Anal. Appl., 254 (2001), 433-463. doi: 10.1006/jmaa.2000.7182
|
| [34] |
S. Ruan, Absolute stability, conditional stability and bifurcation in Kolmogorov-type predator-prey systems with discrete delays, Q. Appl. Math., 59 (2001), 159-173. doi: 10.1090/qam/1811101
|
| [35] | J. Wu, Theory and applications of partial functional differential equations, Springer, New York, 1996. |
| [36] | B. D. Hassard, N. D. Kazarinoff, Y. Wan, Theory and applications of Hopf bifurcation, Cambridge University Press, Cambridge, 1981. |
| [37] |
R. Han, B. Dai, L. Wang, Delay induced spatiotemporal patterns in a diffusive intraguild predation model with Beddington-DeAngelis functional response, Math. Biosci. Eng., 15 (2018), 595-627. doi: 10.3934/mbe.2018027
|
| [38] |
S. Chen, J. Shi, Global dynamics of the diffusive Lotka-Volterra competition model with stage structure, Calculus Var. Partial Differ. Equations, 59 (2020), 1-19. doi: 10.1007/s00526-019-1640-y
|
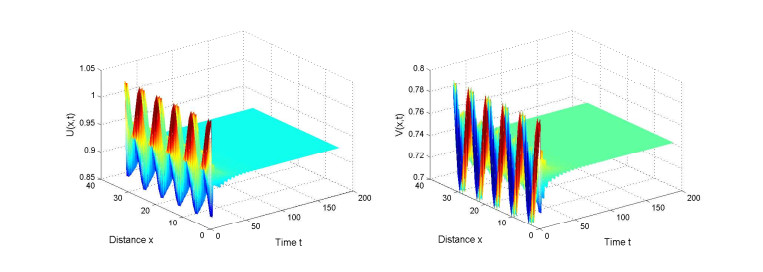
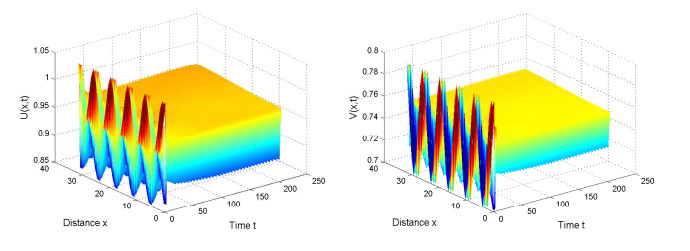
Figures(2)

Changyong Xu, Qiang Li, Tonghua Zhang, Sanling Yuan. Stability and Hopf bifurcation for a delayed diffusive competition model with saturation effect[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 8037-8051. doi: 10.3934/mbe.2020407







 DownLoad:
DownLoad: