Citation: Nao Fukuwada, Miki Kanno, Satomi Yoshida, Kenjiro Seki. Gαq protein signaling in the bed nucleus of the stria terminalis regulate the lipopolysaccharide-induced despair-like behavior in mice[J]. AIMS Neuroscience, 2020, 7(4): 438-458. doi: 10.3934/Neuroscience.2020027

| [1] |
Tiller JW (2013) Depression and anxiety. Med J Aust 199: S28-S31. doi: 10.5694/mja12.10628

|
| [2] |
Choi KW, Kim YK, Jeon HJ (2020) Comorbid Anxiety and Depression: Clinical and Conceptual Consideration and Transdiagnostic Treatment. Adv Exp Med Biol 1191: 219-235. doi: 10.1007/978-981-32-9705-0_14
|
| [3] |
Azad SC, Eder M, Marsicano G, et al. (2003) Activation of the cannabinoid receptor type 1 decreases glutamatergic and GABAergic synaptic transmission in the lateral amygdala of the mouse. Learn Mem 10: 116-128. doi: 10.1101/lm.53303
|
| [4] | Clauss J (2019) Extending the neurocircuitry of behavioural inhibition: a role for the bed nucleus of the stria terminalis in risk for anxiety disorders. Gen Psychiatr 32: 309-315. |
| [5] |
Knight LK, Depue BE (2019) New Frontiers in Anxiety Research: The Translational Potential of the Bed Nucleus of the Stria Terminalis. Front Psychiatry 10: 510. doi: 10.3389/fpsyt.2019.00510
|
| [6] |
Gungor NZ, Paré D (2016) Functional Heterogeneity in the Bed Nucleus of the Stria Terminalis. J Neurosci 36: 8038-8049. doi: 10.1523/JNEUROSCI.0856-16.2016
|
| [7] |
Blomstedt P, Naesström M, Bodlund O (2017) Deep brain stimulation in the bed nucleus of the stria terminalis and medial forebrain bundle in a patient with major depressive disorder and anorexia nervosa. Clin Case Rep 5: 679-684. doi: 10.1002/ccr3.856
|
| [8] |
Drobisz D, Damborská A (2019) Deep brain stimulation targets for treating depression. Behav Brain Res 359: 266-273. doi: 10.1016/j.bbr.2018.11.004
|
| [9] |
Luyck K, Tambuyzer T, Deprez M, et al. (2017) Electrical stimulation of the bed nucleus of the stria terminalis reduces anxiety in a rat model. Transl Psychiatry 7: e1033. doi: 10.1038/tp.2017.2
|
| [10] |
Pezük P, Göz D, Aksoy A, et al. (2006) BNST lesions aggravate behavioral despair but do not impair navigational learning in rats. Brain Res Bull 69: 416-421. doi: 10.1016/j.brainresbull.2006.02.008
|
| [11] |
Schulz D, Canbeyli RS (2000) Lesion of the bed nucleus of the stria terminalis enhances learned despair. Brain Res Bull 52: 83-87. doi: 10.1016/S0361-9230(00)00235-5
|
| [12] |
Crestani CC, Alves FH, Correa FM, et al. (2010) Acute reversible inactivation of the bed nucleus of stria terminalis induces antidepressant-like effect in the rat forced swimming test. Behav Brain Funct 6: 30. doi: 10.1186/1744-9081-6-30
|
| [13] |
Resstel LB, Alves FH, Reis DG, et al. (2008) Anxiolytic-like effects induced by acute reversible inactivation of the bed nucleus of stria terminalis. Neuroscience 154: 869-876. doi: 10.1016/j.neuroscience.2008.04.007
|
| [14] |
Remus JL, Dantzer R (2016) Inflammation Models of Depression in Rodents: Relevance to Psychotropic Drug Discovery. Int J Neuropsychopharmacol 19. doi: 10.1093/ijnp/pyw028
|
| [15] |
Bienkowski MS, Rinaman L (2011) Immune challenge activates neural inputs to the ventrolateral bed nucleus of the stria terminalis. Physiol Behav 104: 257-265. doi: 10.1016/j.physbeh.2011.03.006
|
| [16] | Sekio M, Seki K (2014) Lipopolysaccharide-induced depressive-like behavior is associated with α1-adrenoceptor dependent downregulation of the membrane GluR1 subunit in the mouse medial prefrontal cortex and ventral tegmental area. Int J Neuropsychopharmacol 18. |
| [17] |
Ballesteros-Zebadua P, Manjarrez-Marmolejo J, Franco-Perez J (2013) Chronic paroxetine treatment: effects on other non-serotonergic neurotransmitter systems. CNS Neurol Disord Drug Targets 12: 1226-1232. doi: 10.2174/18715273113126660182
|
| [18] |
Nalepa I, Vetulani J (1993) Enhancement of the responsiveness of cortical adrenergic receptors by chronic administration of the 5-hydroxytryptamine uptake inhibitor citalopram. J Neurochem 60: 2029-2035. doi: 10.1111/j.1471-4159.1993.tb03487.x
|
| [19] |
Vicente MA, Zangrossi H (2014) Involvement of 5-HT2C and 5-HT1A receptors of the basolateral nucleus of the amygdala in the anxiolytic effect of chronic antidepressant treatment. Neuropharmacology 79: 127-135. doi: 10.1016/j.neuropharm.2013.11.007
|
| [20] |
Azevedo H, Ferreira M, Mascarello A, et al. (2020) Brain-wide mapping of c-fos expression in the single prolonged stress model and the effects of pretreatment with ACH-000029 or prazosin. Neurobiol Stress 13: 100226. doi: 10.1016/j.ynstr.2020.100226
|
| [21] |
McElligott ZA, Klug JR, Nobis WP, et al. (2010) Distinct forms of Gq-receptor-dependent plasticity of excitatory transmission in the BNST are differentially affected by stress. Proc Natl Acad Sci USA 107: 2271-2276. doi: 10.1073/pnas.0905568107
|
| [22] |
Itoi K, Sugimoto N (2010) The brainstem noradrenergic systems in stress, anxiety and depression. J Neuroendocrinol 22: 355-361. doi: 10.1111/j.1365-2826.2010.01988.x
|
| [23] |
Itoi K (2008) Ablation of the central noradrenergic neurons for unraveling their roles in stress and anxiety. Ann N Y Acad Sci 1129: 47-54. doi: 10.1196/annals.1417.012
|
| [24] |
Forray MI, Gysling K, Andrés ME, et al. (2000) Medullary noradrenergic neurons projecting to the bed nucleus of the stria terminalis express mRNA for the NMDA-NR1 receptor. Brain Res Bull 52: 163-169. doi: 10.1016/S0361-9230(00)00229-X
|
| [25] |
Qiao H, Li MX, Xu C, et al. (2016) Dendritic Spines in Depression: What We Learned from Animal Models. Neural Plast 2016: 8056370. doi: 10.1155/2016/8056370
|
| [26] | Wright A, Vissel B (2012) The essential role of AMPA receptor GluR2 subunit RNA editing in the normal and diseased brain. Front Mol Neurosci 5: 34. |
| [27] |
Yashiro S, Seki K (2017) Association of social defeat stress-induced anhedonia-like symptoms with mGluR1-dependent decrease in membrane-bound AMPA-GluR1 in the mouse ventral midbrain. Stress 20: 404-418. doi: 10.1080/10253890.2017.1336534
|
| [28] |
Blakemore C, MaCarthur Clark J, Nevalainen T, et al. (2012) Implementing the 3Rs in neuroscience research: a reasoned approach. Neuron 75: 948-950. doi: 10.1016/j.neuron.2012.09.001
|
| [29] | Zhang B, Wang PP, Hu KL, et al. (2019) Antidepressant-Like Effect and Mechanism of Action of Honokiol on the Mouse Lipopolysaccharide (LPS) Depression Model. Molecules 24. |
| [30] |
O'Connor JC, Lawson MA, André C, et al. (2009) Lipopolysaccharide-induced depressive-like behavior is mediated by indoleamine 2,3-dioxygenase activation in mice. Mol Psychiatry 14: 511-522. doi: 10.1038/sj.mp.4002148
|
| [31] |
Walker AK, Wing EE, Banks WA, et al. (2019) Leucine competes with kynurenine for blood-to-brain transport and prevents lipopolysaccharide-induced depression-like behavior in mice. Mol Psychiatry 24: 1523-1532. doi: 10.1038/s41380-018-0076-7
|
| [32] |
Kurosawa N, Shimizu K, Seki K (2016) The development of depression-like behavior is consolidated by IL-6-induced activation of locus coeruleus neurons and IL-1β-induced elevated leptin levels in mice. Psychopharmacology (Berl) 233: 1725-1737. doi: 10.1007/s00213-015-4084-x
|
| [33] |
Shimizu K, Kurosawa N, Seki K (2016) The role of the AMPA receptor and 5-HT(3) receptor on aggressive behavior and depressive-like symptoms in chronic social isolation-reared mice. Physiol Behav 153: 70-83. doi: 10.1016/j.physbeh.2015.10.026
|
| [34] | Franklin K, Paxinos G (2008) The Coronal Plates and Diagrams. The Mouse Brain in Stereotaxic Coordinates, Compact Academic Press, 1-256. |
| [35] |
Williams RW (2000) Mapping genes that modulate mouse brain development: a quantitative genetic approach. Results Probl Cell Differ 30: 21-49. doi: 10.1007/978-3-540-48002-0_2
|
| [36] |
James TA, Starr MS (1978) Effects of the rate and volume of injection on the pharmacological response elicited by intranigral microapplication of drugs in the rat. J Pharmacol Methods 1: 197-202. doi: 10.1016/0160-5402(78)90049-9
|
| [37] |
Yang L, Shi LJ, Yu J, et al. (2016) Activation of protein kinase A in the amygdala modulates anxiety-like behaviors in social defeat exposed mice. Mol Brain 9: 3. doi: 10.1186/s13041-015-0181-3
|
| [38] | Mul JD, Zheng J, Goodyear LJ (2016) Validity Assessment of 5 Day Repeated Forced-Swim Stress to Model Human Depression in Young-Adult C57BL/6J and BALB/cJ Mice. eNeuro 3: 1-9. |
| [39] |
Kanda Y (2013) Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant 48: 452-458. doi: 10.1038/bmt.2012.244
|
| [40] |
Pelloux Y, Hagues G, Costentin J, et al. (2005) Helplessness in the tail suspension test is associated with an increase in ethanol intake and its rewarding effect in female mice. Alcohol Clin Exp Res 29: 378-388. doi: 10.1097/01.ALC.0000156123.10298.FA
|
| [41] |
Perona MT, Waters S, Hall FS, et al. (2008) Animal models of depression in dopamine, serotonin, and norepinephrine transporter knockout mice: prominent effects of dopamine transporter deletions. Behav Pharmacol 19: 566-574. doi: 10.1097/FBP.0b013e32830cd80f
|
| [42] |
Galeotti N, Ghelardini C (2011) Antidepressant phenotype by inhibiting the phospholipase Cβ(1)—protein kinase Cγ pathway in the forced swim test. Neuropharmacology 60: 937-943. doi: 10.1016/j.neuropharm.2011.01.037
|
| [43] |
Chauhan VP (1990) Phosphatidylinositol 4,5-bisphosphate stimulates protein kinase C-mediated phosphorylation of soluble brain proteins. Inhibition by neomycin. FEBS Lett 272: 99-102. doi: 10.1016/0014-5793(90)80457-T
|
| [44] |
Descalzi G, Li XY, Chen T, et al. (2012) Rapid synaptic potentiation within the anterior cingulate cortex mediates trace fear learning. Mol Brain 5: 6. doi: 10.1186/1756-6606-5-6
|
| [45] |
Kodani S, Soya S, Sakurai T (2017) Excitation of GABAergic Neurons in the Bed Nucleus of the Stria Terminalis Triggers Immediate Transition from Non-Rapid Eye Movement Sleep to Wakefulness in Mice. J Neurosci 37: 7164-7176. doi: 10.1523/JNEUROSCI.0245-17.2017
|
| [46] |
Woulfe JM, Hrycyshyn AW, Flumerfelt BA (1988) Collateral axonal projections from the A1 noradrenergic cell group to the paraventricular nucleus and bed nucleus of the stria terminalis in the rat. Exp Neurol 102: 121-124. doi: 10.1016/0014-4886(88)90084-2
|
| [47] |
Mejías-Aponte CA, Drouin C, Aston-Jones G (2009) Adrenergic and noradrenergic innervation of the midbrain ventral tegmental area and retrorubral field: prominent inputs from medullary homeostatic centers. J Neurosci 29: 3613-3626. doi: 10.1523/JNEUROSCI.4632-08.2009
|
| [48] |
Dumont EC, Williams JT (2004) Noradrenaline triggers GABAA inhibition of bed nucleus of the stria terminalis neurons projecting to the ventral tegmental area. J Neurosci 24: 8198-8204. doi: 10.1523/JNEUROSCI.0425-04.2004
|
| [49] |
Velásquez-Martínez MC, Vázquez-Torres R, Rojas LV, et al. (2015) Alpha-1 adrenoreceptors modulate GABA release onto ventral tegmental area dopamine neurons. Neuropharmacology 88: 110-121. doi: 10.1016/j.neuropharm.2014.09.002
|
| [50] |
Paladini CA, Williams JT (2004) Noradrenergic inhibition of midbrain dopamine neurons. J Neurosci 24: 4568-4575. doi: 10.1523/JNEUROSCI.5735-03.2004
|
| [51] |
Mejias-Aponte CA (2016) Specificity and impact of adrenergic projections to the midbrain dopamine system. Brain Res 1641: 258-273. doi: 10.1016/j.brainres.2016.01.036
|
| [52] |
Dyer-Reaves K, Goodman AM, Nelson AR, et al. (2019) Alpha1-Adrenergic Receptor Mediated Long-Term Depression at CA3-CA1 Synapses Can Be Induced via Accumulation of Endogenous Norepinephrine and Is Preserved Following Noradrenergic Denervation. Front Synaptic Neurosci 11: 27. doi: 10.3389/fnsyn.2019.00027
|
| [53] |
Scheiderer CL, Dobrunz LE, McMahon LL (2004) Novel form of long-term synaptic depression in rat hippocampus induced by activation of alpha 1 adrenergic receptors. J Neurophysiol 91: 1071-1077. doi: 10.1152/jn.00420.2003
|
| [54] |
Choi SY, Chang J, Jiang B, et al. (2005) Multiple receptors coupled to phospholipase C gate long-term depression in visual cortex. J Neurosci 25: 11433-11443. doi: 10.1523/JNEUROSCI.4084-05.2005
|
| [55] |
Kirkwood A, Rozas C, Kirkwood J, et al. (1999) Modulation of long-term synaptic depression in visual cortex by acetylcholine and norepinephrine. J Neurosci 19: 1599-1609. doi: 10.1523/JNEUROSCI.19-05-01599.1999
|
| [56] |
Jo JH, Park EJ, Lee JK, et al. (2001) Lipopolysaccharide inhibits induction of long-term potentiation and depression in the rat hippocampal CA1 area. Eur J Pharmacol 422: 69-76. doi: 10.1016/S0014-2999(01)01075-5
|
| [57] |
Sagar SM, Price KJ, Kasting NW, et al. (1995) Anatomic patterns of Fos immunostaining in rat brain following systemic endotoxin administration. Brain Res Bull 36: 381-392. doi: 10.1016/0361-9230(94)00217-O
|
| [58] |
Gaykema RP, Daniels TE, Shapiro NJ, et al. (2009) Immune challenge and satiety-related activation of both distinct and overlapping neuronal populations in the brainstem indicate parallel pathways for viscerosensory signaling. Brain Res 1294: 61-79. doi: 10.1016/j.brainres.2009.07.076
|
| [59] |
Tanaka M, Yoshida M, Emoto H, et al. (2000) Noradrenaline systems in the hypothalamus, amygdala and locus coeruleus are involved in the provocation of anxiety: basic studies. Eur J Pharmacol 405: 397-406. doi: 10.1016/S0014-2999(00)00569-0
|
| [60] |
Southwick SM, Bremner JD, Rasmusson A, et al. (1999) Role of norepinephrine in the pathophysiology and treatment of posttraumatic stress disorder. Biol Psychiatry 46: 1192-1204. doi: 10.1016/S0006-3223(99)00219-X
|
| [61] |
Perahia DG, Quail D, Desaiah D, et al. (2008) Switching to duloxetine from selective serotonin reuptake inhibitor antidepressants: a multicenter trial comparing 2 switching techniques. J Clin Psychiatry 69: 95-105. doi: 10.4088/JCP.v69n0113
|
| [62] |
Nutt DJ (2002) The neuropharmacology of serotonin and noradrenaline in depression. Int Clin Psychopharmacol 17 Suppl 1: S1-S12. doi: 10.1097/00004850-200206001-00002
|
| [63] |
Sethy VH, Day JS, Cooper MM (1988) Dose-dependent down-regulation of beta-adrenergic receptors after chronic intravenous infusion of antidepressants. Prog Neuropsychopharmacol Biol Psychiatry 12: 673-682. doi: 10.1016/0278-5846(88)90011-5
|
| [64] |
Kreiner G, Zelek-Molik A, Kowalska M, et al. (2011) Effects of the noradrenergic neurotoxin DSP-4 on the expression of α1-adrenoceptor subtypes after antidepressant treatment. Pharmacol Rep 63: 1349-1358. doi: 10.1016/S1734-1140(11)70699-5
|
| [65] |
Heisler LK, Zhou L, Bajwa P, et al. (2007) Serotonin 5-HT(2C) receptors regulate anxiety-like behavior. Genes Brain Behav 6: 491-496. doi: 10.1111/j.1601-183X.2007.00316.x
|
| [66] |
Lebow MA, Chen A (2016) Overshadowed by the amygdala: the bed nucleus of the stria terminalis emerges as key to psychiatric disorders. Mol Psychiatry 21: 450-463. doi: 10.1038/mp.2016.1
|
| [67] |
Kim SY, Adhikari A, Lee SY, et al. (2013) Diverging neural pathways assemble a behavioural state from separable features in anxiety. Nature 496: 219-223. doi: 10.1038/nature12018
|
| [68] |
Dong HW, Petrovich GD, Watts AG, et al. (2001) Basic organization of projections from the oval and fusiform nuclei of the bed nuclei of the stria terminalis in adult rat brain. J Comp Neurol 436: 430-455. doi: 10.1002/cne.1079
|
| [69] |
Choi DC, Furay AR, Evanson NK, et al. (2007) Bed nucleus of the stria terminalis subregions differentially regulate hypothalamic-pituitary-adrenal axis activity: implications for the integration of limbic inputs. J Neurosci 27: 2025-2034. doi: 10.1523/JNEUROSCI.4301-06.2007
|
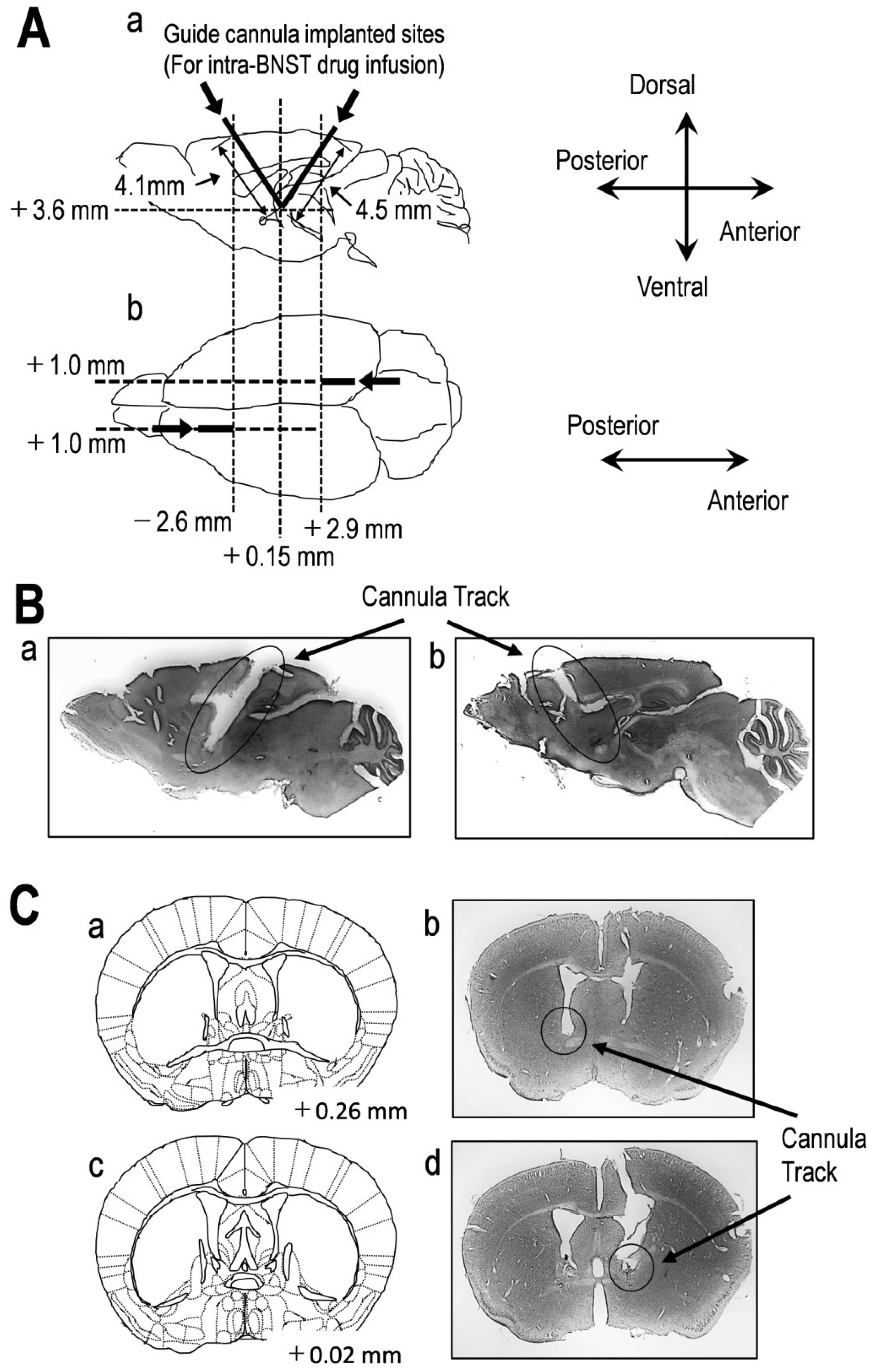
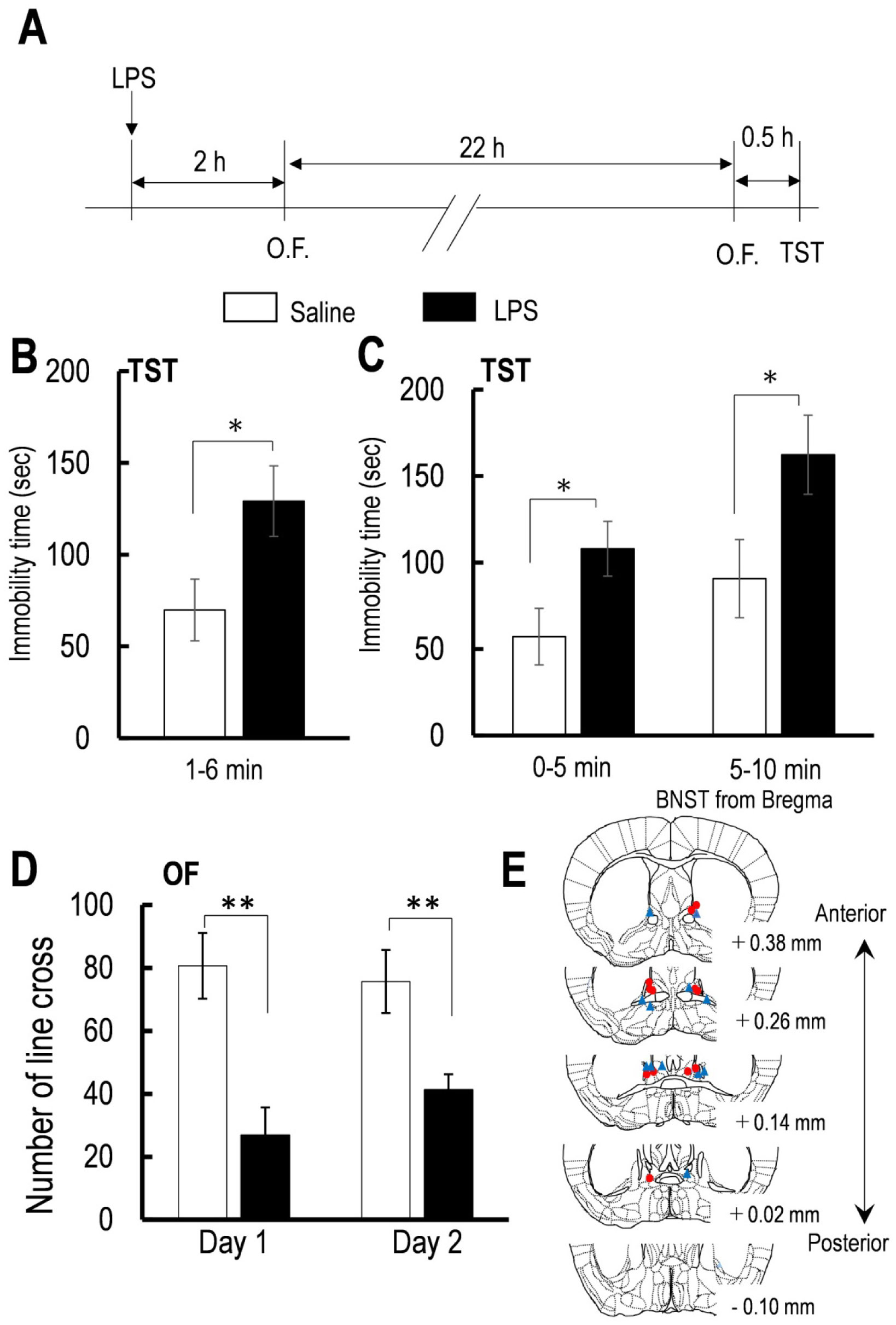
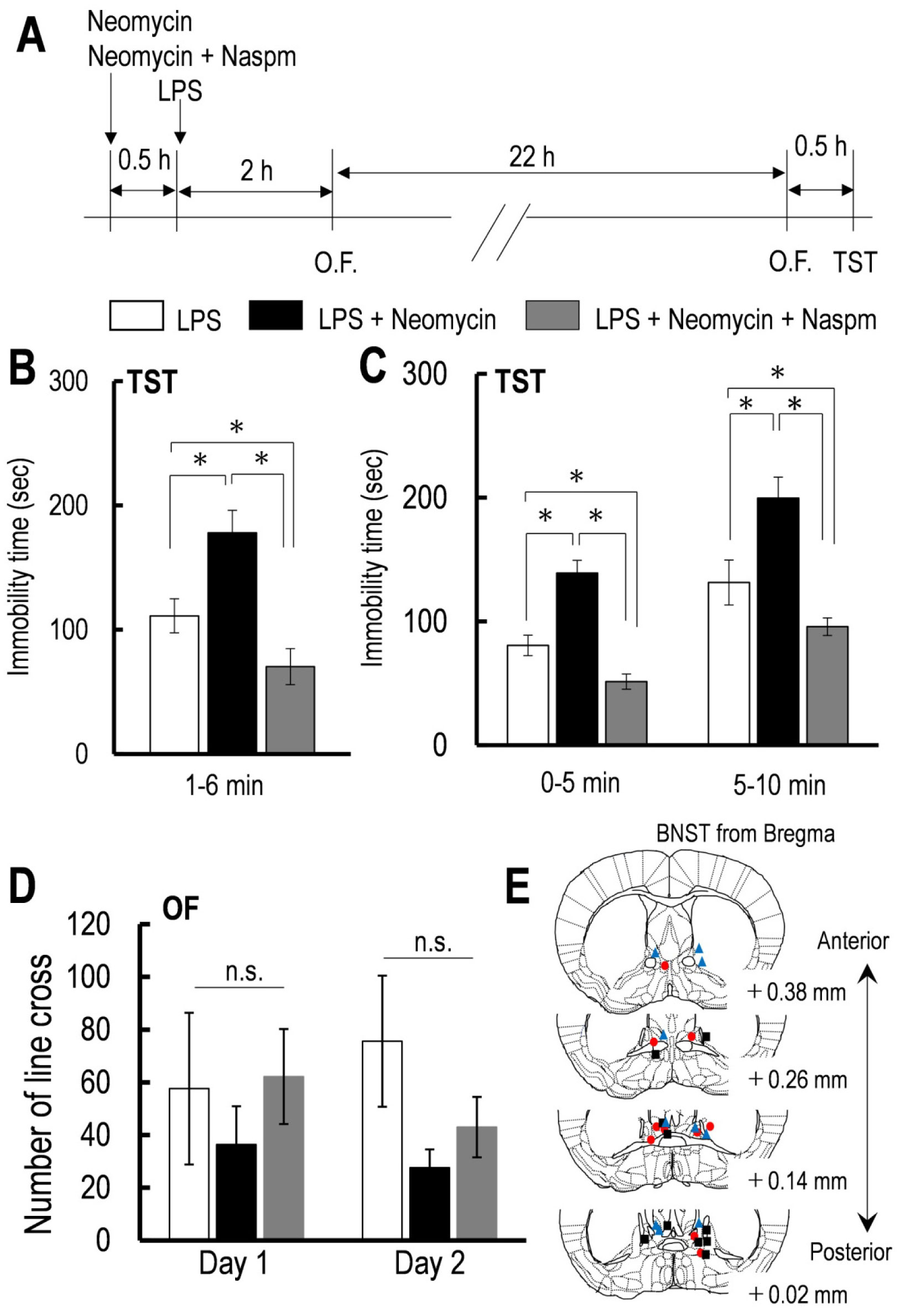

Figures(4)

Nao Fukuwada, Miki Kanno, Satomi Yoshida, Kenjiro Seki. Gαq protein signaling in the bed nucleus of the stria terminalis regulate the lipopolysaccharide-induced despair-like behavior in mice[J]. AIMS Neuroscience, 2020, 7(4): 438-458. doi: 10.3934/Neuroscience.2020027







 DownLoad:
DownLoad: