Citation: Onyekachi Juliet Okpasuo, Ifeanyi Oscar Aguzie, Anunobi Toochukwu Joy, Fabian C Okafor. Risk assessment of waterborne infections in Enugu State, Nigeria: Implications of household water choices, knowledge, and practices[J]. AIMS Public Health, 2020, 7(3): 634-649. doi: 10.3934/publichealth.2020050

| [1] | Ayoade AA, Sikiru S, Okanlawon PO (2017) Assessment of Water Provision and Associated Risks Among Children in Abeokuta Peri-Urban, Ogun State, Southwestern Nigeria: The Gender Implications. wH2O: J Gender Water 4: 9. |
| [2] | World Health Organisation (2010) Progress on sanitation and drinking-water: Joint Monitoring Programme 2010 update. Available from: https://www.who.int/water_sanitation_health/publications/9789241563956/en. |
| [3] | United Nations Children Education Fund (UNICEF)/World Health Organization (WHO) (2012) Progress on drinking water and sanitation-2012 update, USA. Available from: https://www.unicef.org/media/files/JMPreport2012.pdf. |
| [4] | World Health Organization (2000) Global water supply and sanitation assessment report. Available from: https://www.who.int/water_sanitation_health/monitoring/jmp2000/en/. |
| [5] | World Health Organization (2012) Burden of Diseases and Cost-Effectiveness Estimates. Available from: https://www.en.wikipedia.org/wiki/waterborne-diseases. |
| [6] | United Nations Children's Fund (UNICEF)/World Health Organization (WHO) (2009) Diarrhoea: Why Children are still dying and what can be done. Available from: https://www.who.int/maternal_child_adolescent/document/9789241598415/2n/. |
| [7] |
Johnson JYM, Thomas JE, Graham TA, et al. (2003) Prevalence of Escherichia coli 0157: H7 and Salmonella spp, in surface waters of Southern Alberts and its relation to manure source. Can J Microbiol 49: 326-335. doi: 10.1139/w03-046

|
| [8] | Obiri-Danso K, Adjei B, Stanley KN, et al. (2009) Microbiological quality and metal levels in wells and boreholes water in some peri-urban communities in Kumasi, Ghana. Afr J Environ Sci Technol 3: 059-066. |
| [9] | CNN Wire Staff (2010) Waterborne diseases outbreaks in developing countries. Available from: http://edition.cnn.com/2010/WORLD/africa/. |
| [10] |
Armah FA, Ekumah B, Yawson DO, et al. (2018) Access to improved water and sanitation in sub-Saharan Africa in a quarter century. Heliyon 4: e00931. doi: 10.1016/j.heliyon.2018.e00931
|
| [11] | Malik A, Yasar A, Tabinda A, et al. (2012) Water-borne diseases, cost of illness and willingness to pay for diseases interventions in rural communities of developing countries. Iran J Public Health 41: 39-49. |
| [12] | WASH-Plus (2010) WASH related diseases. Available from: http://washalerts.wordpress.com/author/envhealth/page/22/. |
| [13] | Monique U (2012) Microbiological drinking water quality and prevalence of waterborne diseases in Masaka, Rwanda Durban, South Africa: Thesis, Durban University of Technology. |
| [14] | Fletcher SM, Mary-Louise M, John TE (2013) Prevalence of gastrointestinal pathogens in developed and developing countries: systematic review and meta-analysis. J Public Health Resour 2: 42-53. |
| [15] |
Pandey PK, Kass PH, Soupir ML, et al. (2014) Contamination of water resources by pathogenic bacteria. AMB Express 4: 51. doi: 10.1186/s13568-014-0051-x
|
| [16] | Okpasuo OJ, Okafor FC, Aguzie I, et al. (2019) Spatiotemporal trend of waterborne disease in Enugu Urban, Nigeria: A retrospective study. Int J Trop Dis Health 38: 1-13. |
| [17] | Olajuyigbe AE, Alinaitwe P, Adegboyega SA, et al. (2012) Spatial Analysis of Factors Responsible for Incidence of Water Borne Diseases in Ile-Ife, Nigeria. J Sustainable Soc 1: 96-113. |
| [18] | Akabuike BO (1990) A Short Text on Geography Enugu: Pegant Ventures. |
| [19] | Thomas B (2017) City population. Available from: http://www.citypopulation.info/php/nigeria-admin.php?adm2id=NGA014005. |
| [20] |
Ezenwaji EE, Anyadike RNC, Igu NI (2014) Optimal allocation of public water supply to the urban sectors of Enugu, Nigeria: a linear programming approach. Appl Water Sci 4: 73-78. doi: 10.1007/s13201-013-0131-0
|
| [21] | Vandepitte J, Verhaegen J, Engbaek K, et al. (2003) Basic laboratory procedures in clinical bacteriology Geneva: World Health Organization. |
| [22] |
Cheesbrough M (2006) District laboratory practice in tropical countries part 2 New York: Cambridge University Press. doi: 10.1017/CBO9780511543470
|
| [23] |
Odeyemi OA (2015) Bacteriological safety of packaged drinking water sold in Nigeria: public health implications. Springer plus 4: 642. doi: 10.1186/s40064-015-1447-z
|
| [24] | Olajuyigbe AE (2010) Some factors impacting on the quantity of water used by households in a rapidly urbanizing State capital in South Western Nigeria. J Sustainable Dev Afr 12: 322-337. |
| [25] |
Dalhat MM, Isa AN, Nguku P, et al. (2014) Descriptive characterization of the 2010 cholera outbreak in Nigeria. BMC Public Health 14: 1167. doi: 10.1186/1471-2458-14-1167
|
| [26] | Sule IB, Yahaya M, Aisha AA, et al. (2014) Descriptive epidemiology of a cholera outbreak in Kaduna State, Northwest Nigeria. Pan Afr Med J 27: 172. |
| [27] | Raji MO, Ibrahim Y (2011) Prevalence of waterborne infections in Northwest Nigeria: A retrospective study. J Public Health Epidemiol 3: 382-385. |
| [28] | (2014) United State Agency for International Development (USAID)Nigeria Water and Sanitation Profile. London, United Kingdom: University Press. |
| [29] | WHO (2014) Water Quality and Health. Drinking water chlorination-A review of disinfection practices and issues. Available from: http://www.waterandhealth.org/drinkingwater/wp.html. |
| [30] |
Das JK, Salam RA, Bhutta ZA (2014) Global burden of childhood diarrhea and interventions. Curr Opin Infect Dis 27: 451-458. doi: 10.1097/QCO.0000000000000096
|
| [31] |
Yang K, LeJeune J, Alsdorf D, et al. (2012) Global distribution of outbreaks of water-associated infectious diseases. PLoS Negl Trop Dis 6: e1483. doi: 10.1371/journal.pntd.0001483
|
| [32] |
Hindman PT (2002) Household choice of drinking-water source in the Philippines. Asian Econ J 16: 303-316. doi: 10.1111/1467-8381.t01-1-00154
|
| [33] | Onjala J, Ndiritu SW, Stage J (2013) Risk Perception, Choice of Drinking Water, and Water Treatment. Environ Dev. 13-10. |
| [34] |
Rosa G, Miller L, Clasen T (2010) Microbiological Effectiveness of Disinfecting Water by Boiling in Rural Guatemala. Am J Trop Med Hyg 82: 473-477. doi: 10.4269/ajtmh.2010.09-0320
|
| [35] |
Kioko KJ, Obiri JF (2012) Household attitudes and knowledge on drinking water enhance water hazards in peri-urban communities in Western Kenya. Jàmbá J Disaster Risk Stud 4: 49-54. doi: 10.4102/jamba.v4i1.49
|
| [36] | Dunker L (2001) The KAP Tool for Hygiene. A Manual on: Knowledge, Attitudes and Practices in the Rural Areas of South Africa. WRC Report No.TT 144/00, Water Research Commission, Pretoria, South Africa . |
| [37] | Nala NP, Jagals P, Joubert G (2003) The effect of a water-hygiene educational programme on the microbiological quality of container-stored water in households. Water SA 29: 171. |
| [38] | Lantagne D, Quick R, Mintz E (2006) Household water treatment and safe storage options in developing countries: a review of current implementation practices Washington DC: Woodrow Wilson International Center. |
| [39] | Center for Disease Control (2010) Global Water Sanitation and Hygiene. Available from: http://www.cdc.gov/healthywater/global/index.html. |
| [40] | Pruss-Ustun A, Bos R, Gore F, et al. (2012) Safe Water, Better Health: Cost, Benefits and Sustainability of Interventions to Protect and Promote Health Geneva, Switzerland: World Health Organization. |
 publichealth-07-03-050-s001.pdf publichealth-07-03-050-s001.pdf |
 |
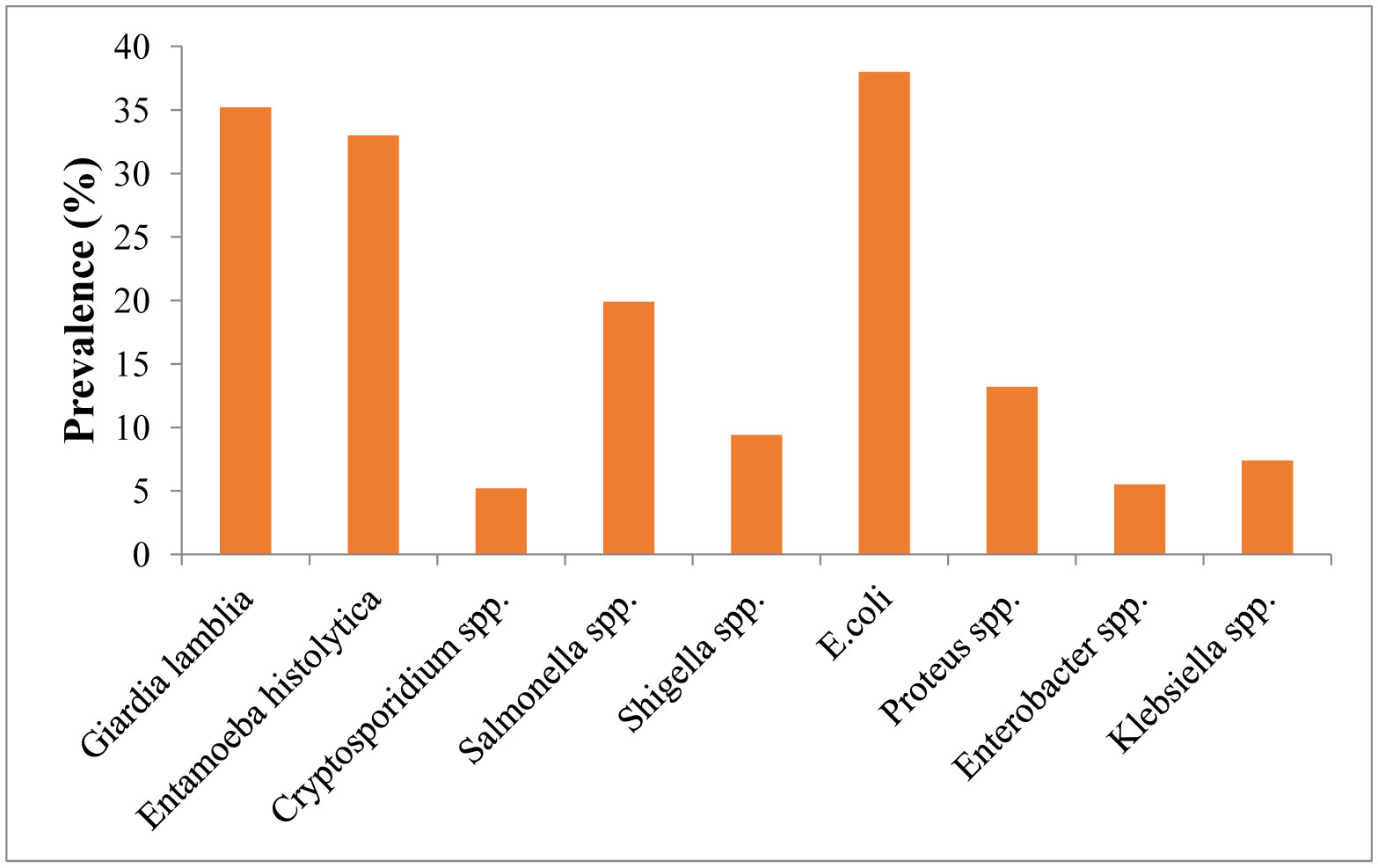
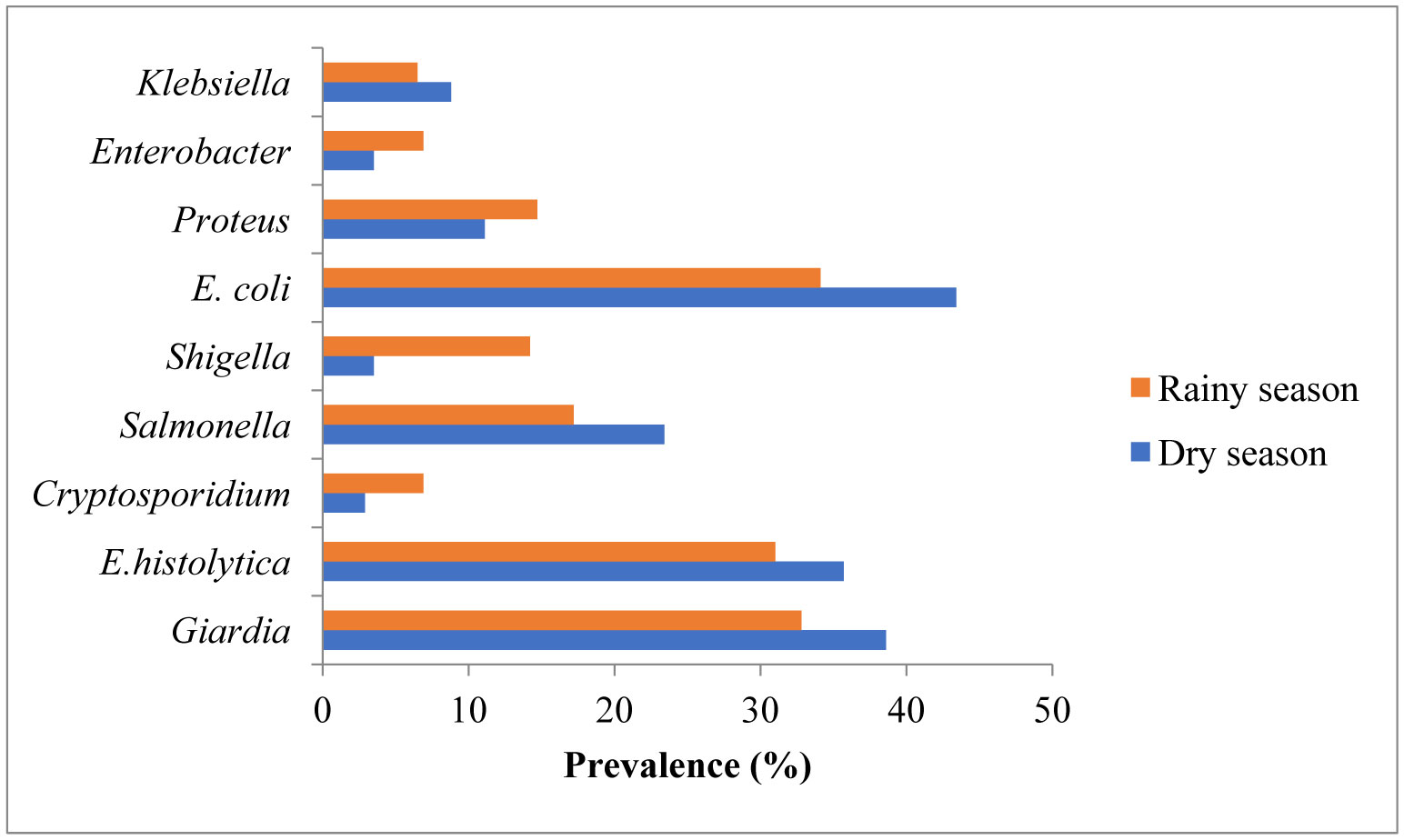
Figures(2) / Tables(4)

Onyekachi Juliet Okpasuo, Ifeanyi Oscar Aguzie, Anunobi Toochukwu Joy, Fabian C Okafor. Risk assessment of waterborne infections in Enugu State, Nigeria: Implications of household water choices, knowledge, and practices[J]. AIMS Public Health, 2020, 7(3): 634-649. doi: 10.3934/publichealth.2020050







 DownLoad:
DownLoad: