Citation: Cassandra A DeMarshall, Abhirup Sarkar, Robert G Nagele. Serum Autoantibodies as Biomarkers for Parkinsons Disease: Background and Utility[J]. AIMS Medical Science, 2015, 2(4): 316-327. doi: 10.3934/medsci.2015.4.316

| [1] |
Jankovic J (2008) Parkinson's disease: clinical features and diagnosis. J Neurol Neurosurg Psychiatry 79: 368-376. doi: 10.1136/jnnp.2007.131045

|
| [2] |
de Lau LM, Breteler MM (2006) Epidemiology of Parkinson's disease. Lancet Neurol 5: 525-535. doi: 10.1016/S1474-4422(06)70471-9
|
| [3] |
Davie CA (2008) A review of Parkinson's disease. Br Med Bull 86: 109-127. doi: 10.1093/bmb/ldn013
|
| [4] |
Aarsland D, Beyer MK, Kurz MW (2008) Dementia in Parkinson's disease. Curr Opin Neurol 21: 676-682. doi: 10.1097/WCO.0b013e3283168df0
|
| [5] |
Adler CH, Dugger BN, Hinni ML, et al. (2014) Submandibular gland needle biopsy for the diagnosis of Parkinson disease. Neurology 82: 858-864. doi: 10.1212/WNL.0000000000000204
|
| [6] | Aerts MB, Esselink RA, Abdo WF, et al. (2012) CSF alpha-synuclein does not differentiate between parkinsonian disorders. Neurobiol Aging 33: 430 e431-433. |
| [7] |
Mollenhauer B, Cullen V, Kahn I, et al. (2008) Direct quantification of CSF alpha-synuclein by ELISA and first cross-sectional study in patients with neurodegeneration. Exp Neurol 213: 315-325. doi: 10.1016/j.expneurol.2008.06.004
|
| [8] | van Dijk KD, Bidinosti M, Weiss A, et al. (2014) Reduced alpha-synuclein levels in cerebrospinal fluid in Parkinson's disease are unrelated to clinical and imaging measures of disease severity. Eur J Neurol 21: 388-394. |
| [9] |
Gerlach M, Maetzler W, Broich K, et al. (2012) Biomarker candidates of neurodegeneration in Parkinson's disease for the evaluation of disease-modifying therapeutics. J Neural Transm 119: 39-52. doi: 10.1007/s00702-011-0682-x
|
| [10] | Gao L, Chen H, Li X, et al. (2015) The diagnostic value of minor salivary gland biopsy in clinically diagnosed patients with Parkinson's disease: comparison with DAT PET scans. Neurol Sci. |
| [11] |
Folgoas E, Lebouvier T, Leclair-Visonneau L, et al. (2013) Diagnostic value of minor salivary glands biopsy for the detection of Lewy pathology. Neurosci Lett 551: 62-64. doi: 10.1016/j.neulet.2013.07.016
|
| [12] | Agrawal S, Misra R, Aggarwal A (2007) Autoantibodies in rheumatoid arthritis: association with severity of disease in established RA. Clin Rheumatol 26: 201-204. |
| [13] |
Sherer Y, Gorstein A, Fritzler MJ, et al. (2004) Autoantibody explosion in systemic lupus erythematosus: more than 100 different antibodies found in SLE patients. Semin Arthritis Rheum 34: 501-537. doi: 10.1016/j.semarthrit.2004.07.002
|
| [14] |
Diamond B, Huerta PT, Mina-Osorio P, et al. (2009) Losing your nerves? Maybe it's the antibodies. Nat Rev Immunol 9: 449-456. doi: 10.1038/nri2529
|
| [15] |
Tan HT, Low J, Lim SG, et al. (2009) Serum autoantibodies as biomarkers for early cancer detection. FEBS J 276: 6880-6904. doi: 10.1111/j.1742-4658.2009.07396.x
|
| [16] |
Levin EC, Acharya NK, Han M, et al. (2010) Brain-reactive autoantibodies are nearly ubiquitous in human sera and may be linked to pathology in the context of blood-brain barrier breakdown. Brain Res 1345: 221-232. doi: 10.1016/j.brainres.2010.05.038
|
| [17] |
Nagele EP, Han M, Acharya NK, et al. (2013) Natural IgG autoantibodies are abundant and ubiquitous in human sera, and their number is influenced by age, gender, and disease. PLoS One 8: e60726. doi: 10.1371/journal.pone.0060726
|
| [18] |
Lacroix-Desmazes S, Mouthon L, Kaveri SV, et al. (1999) Stability of natural self-reactive antibody repertoires during aging. J Clin Immunol 19: 26-34. doi: 10.1023/A:1020510401233
|
| [19] |
Mirilas P, Fesel C, Guilbert B, et al. (1999) Natural antibodies in childhood: development, individual stability, and injury effect indicate a contribution to immune memory. J Clin Immunol 19: 109-115. doi: 10.1023/A:1020554500266
|
| [20] | Avrameas S (1991) Natural autoantibodies: from 'horror autotoxicus' to 'gnothi seauton'. Immunol Today 12: 154-159. |
| [21] |
Avrameas S, Ternynck T, Tsonis IA, et al. (2007) Naturally occurring B-cell autoreactivity: a critical overview. J Autoimmun 29: 213-218. doi: 10.1016/j.jaut.2007.07.010
|
| [22] | Fearnley JM, Lees AJ (1991) Ageing and Parkinson's disease: substantia nigra regional selectivity. Brain 114 ( Pt 5): 2283-2301. |
| [23] |
Postuma RB, Gagnon JF, Montplaisir J (2010) Clinical prediction of Parkinson's disease: planning for the age of neuroprotection. J Neurol Neurosurg Psychiatry 81: 1008-1013. doi: 10.1136/jnnp.2009.174748
|
| [24] |
Nagele E, Han M, Demarshall C, et al. (2011) Diagnosis of Alzheimer's disease based on disease-specific autoantibody profiles in human sera. PLoS One 6: e23112. doi: 10.1371/journal.pone.0023112
|
| [25] |
Han M, Nagele E, DeMarshall C, et al. (2012) Diagnosis of Parkinson's disease based on disease-specific autoantibody profiles in human sera. PLoS One 7: e32383. doi: 10.1371/journal.pone.0032383
|
| [26] |
DeMarshall CA, Han M, Nagele EP, et al. (2015) Potential utility of autoantibodies as blood-based biomarkers for early detection and diagnosis of Parkinson's disease. Immunol Lett 168: 80-88. doi: 10.1016/j.imlet.2015.09.010
|
| [27] | Rice JS, Kowal C, Volpe BT, et al. (2005) Molecular mimicry: anti-DNA antibodies bind microbial and nonnucleic acid self-antigens. Curr Top Microbiol Immunol 296: 137-151. |
| [28] | Nagele RG, Clifford PM, Siu G, et al. (2011) Brain-reactive autoantibodies prevalent in human sera increase intraneuronal amyloid-beta(1-42) deposition. J Alzheimers Dis 25: 605-622. |
| [29] |
Nath A, Hall E, Tuzova M, et al. (2003) Autoantibodies to amyloid beta-peptide (Abeta) are increased in Alzheimer's disease patients and Abeta antibodies can enhance Abeta neurotoxicity: implications for disease pathogenesis and vaccine development. Neuromolecular Med 3: 29-39. doi: 10.1385/NMM:3:1:29
|
| [30] |
Maftei M, Thurm F, Schnack C, et al. (2013) Increased levels of antigen-bound beta-amyloid autoantibodies in serum and cerebrospinal fluid of Alzheimer's disease patients. PLoS One 8: e68996. doi: 10.1371/journal.pone.0068996
|
| [31] | Maetzler W, Berg D, Synofzik M, et al. (2011) Autoantibodies against amyloid and glial-derived antigens are increased in serum and cerebrospinal fluid of Lewy body-associated dementias. J Alzheimers Dis 26: 171-179. |
| [32] |
Costa A, Bini P, Hamze-Sinno M, et al. (2011) Galanin and alpha-MSH autoantibodies in cerebrospinal fluid of patients with Alzheimer's disease. J Neuroimmunol 240-241: 114-120. doi: 10.1016/j.jneuroim.2011.10.003
|
| [33] |
McIntyre JA, Ramsey CJ, Gitter BD, et al. (2015) Antiphospholipid autoantibodies as blood biomarkers for detection of early stage Alzheimer's disease. Autoimmunity 48: 344-351. doi: 10.3109/08916934.2015.1008464
|
| [34] |
Conradi S, Ronnevi LO (1993) Selective vulnerability of alpha motor neurons in ALS: relation to autoantibodies toward acetylcholinesterase (AChE) in ALS patients. Brain Res Bull 30: 369-371. doi: 10.1016/0361-9230(93)90267-F
|
| [35] |
Tzartos JS, Zisimopoulou P, Rentzos M, et al. (2014) LRP4 antibodies in serum and CSF from amyotrophic lateral sclerosis patients. Ann Clin Transl Neurol 1: 80-87. doi: 10.1002/acn3.26
|
| [36] |
Fialova L, Svarcova J, Bartos A, et al. (2010) Cerebrospinal fluid and serum antibodies against neurofilaments in patients with amyotrophic lateral sclerosis. Eur J Neurol 17: 562-566. doi: 10.1111/j.1468-1331.2009.02853.x
|
| [37] |
Lee DH, Heidecke H, Schroder A, et al. (2014) Increase of angiotensin II type 1 receptor auto-antibodies in Huntington's disease. Mol Neurodegener 9: 49. doi: 10.1186/1750-1326-9-49
|
| [38] |
Pollak TA, McCormack R, Peakman M, et al. (2014) Prevalence of anti-N-methyl-D-aspartate (NMDA) receptor [corrected] antibodies in patients with schizophrenia and related psychoses: a systematic review and meta-analysis. Psychol Med 44: 2475-2487. doi: 10.1017/S003329171300295X
|
| [39] |
Pearlman DM, Najjar S (2014) Meta-analysis of the association between N-methyl-d-aspartate receptor antibodies and schizophrenia, schizoaffective disorder, bipolar disorder, and major depressive disorder. Schizophr Res 157: 249-258. doi: 10.1016/j.schres.2014.05.001
|
| [40] |
Levite M, Ganor Y (2008) Autoantibodies to glutamate receptors can damage the brain in epilepsy, systemic lupus erythematosus and encephalitis. Expert Rev Neurother 8: 1141-1160. doi: 10.1586/14737175.8.7.1141
|
| [41] |
Lee JY, Huerta PT, Zhang J, et al. (2009) Neurotoxic autoantibodies mediate congenital cortical impairment of offspring in maternal lupus. Nat Med 15: 91-96. doi: 10.1038/nm.1892
|
| [42] |
Zimmerman AW, Connors SL, Matteson KJ, et al. (2007) Maternal antibrain antibodies in autism. Brain Behav Immun 21: 351-357. doi: 10.1016/j.bbi.2006.08.005
|
| [43] | Braunschweig D, Ashwood P, Krakowiak P, et al. (2008) Autism: maternally derived antibodies specific for fetal brain proteins. Neurotoxicology 29: 226-231. |
| [44] |
Cabanlit M, Wills S, Goines P, et al. (2007) Brain-specific autoantibodies in the plasma of subjects with autistic spectrum disorder. Ann N Y Acad Sci 1107: 92-103. doi: 10.1196/annals.1381.010
|
| [45] |
Singer HS, Morris CM, Williams PN, et al. (2006) Antibrain antibodies in children with autism and their unaffected siblings. J Neuroimmunol 178: 149-155. doi: 10.1016/j.jneuroim.2006.05.025
|
| [46] | Raad M, Nohra E, Chams N, et al. (2014) Autoantibodies in traumatic brain injury and central nervous system trauma. Neuroscience 281C: 16-23. |
| [47] | Kobeissy F, Moshourab RA (2015) Autoantibodies in CNS Trauma and Neuropsychiatric Disorders: A New Generation of Biomarkers. In: Kobeissy FHP, editor. Brain Neurotrauma: Molecular, Neuropsychological, and Rehabilitation Aspects. Boca Raton (FL). |
| [48] | Zhang Y, Popovich P (2011) Roles of autoantibodies in central nervous system injury. Discov Med 11: 395-402. |
| [49] |
Skoda D, Kranda K, Bojar M, et al. (2006) Antibody formation against beta-tubulin class III in response to brain trauma. Brain Res Bull 68: 213-216. doi: 10.1016/j.brainresbull.2005.05.032
|
| [50] |
Davies AL, Hayes KC, Dekaban GA (2007) Clinical correlates of elevated serum concentrations of cytokines and autoantibodies in patients with spinal cord injury. Arch Phys Med Rehabil 88: 1384-1393. doi: 10.1016/j.apmr.2007.08.004
|
| [51] |
Spillantini MG, Schmidt ML, Lee VM, et al. (1997) Alpha-synuclein in Lewy bodies. Nature 388: 839-840. doi: 10.1038/42166
|
| [52] | Marui W, Iseki E, Kato M, et al. (2004) Pathological entity of dementia with Lewy bodies and its differentiation from Alzheimer's disease. Acta Neuropathol 108: 121-128. |
| [53] |
Ohrfelt A, Grognet P, Andreasen N, et al. (2009) Cerebrospinal fluid alpha-synuclein in neurodegenerative disorders-a marker of synapse loss? Neurosci Lett 450: 332-335. doi: 10.1016/j.neulet.2008.11.015
|
| [54] |
Hall S, Surova Y, Ohrfelt A, et al. (2015) CSF biomarkers and clinical progression of Parkinson disease. Neurology 84: 57-63. doi: 10.1212/WNL.0000000000001098
|
| [55] |
Mondello S, Constantinescu R, Zetterberg H, et al. (2014) CSF alpha-synuclein and UCH-L1 levels in Parkinson's disease and atypical parkinsonian disorders. Parkinsonism Relat Disord 20: 382-387. doi: 10.1016/j.parkreldis.2014.01.011
|
| [56] |
Kruse N, Persson S, Alcolea D, et al. (2015) Validation of a quantitative cerebrospinal fluid alpha-synuclein assay in a European-wide interlaboratory study. Neurobiol Aging 36: 2587-2596. doi: 10.1016/j.neurobiolaging.2015.05.003
|
| [57] |
Koehler NK, Stransky E, Meyer M, et al. (2015) Alpha-synuclein levels in blood plasma decline with healthy aging. PLoS One 10: e0123444. doi: 10.1371/journal.pone.0123444
|
| [58] |
Caranci G, Piscopo P, Rivabene R, et al. (2013) Gender differences in Parkinson's disease: focus on plasma alpha-synuclein. J Neural Transm 120: 1209-1215. doi: 10.1007/s00702-013-0972-6
|
| [59] |
Besong-Agbo D, Wolf E, Jessen F, et al. (2013) Naturally occurring alpha-synuclein autoantibody levels are lower in patients with Parkinson disease. Neurology 80: 169-175. doi: 10.1212/WNL.0b013e31827b90d1
|
| [60] |
Yanamandra K, Gruden MA, Casaite V, et al. (2011) alpha-synuclein reactive antibodies as diagnostic biomarkers in blood sera of Parkinson's disease patients. PLoS One 6: e18513. doi: 10.1371/journal.pone.0018513
|
| [61] |
Smith LM, Schiess MC, Coffey MP, et al. (2012) alpha-Synuclein and anti-alpha-synuclein antibodies in Parkinson's disease, atypical Parkinson syndromes, REM sleep behavior disorder, and healthy controls. PLoS One 7: e52285. doi: 10.1371/journal.pone.0052285
|
| [62] |
Double KL, Rowe DB, Carew-Jones FM, et al. (2009) Anti-melanin antibodies are increased in sera in Parkinson's disease. Exp Neurol 217: 297-301. doi: 10.1016/j.expneurol.2009.03.002
|
| [63] | Zappia M, Crescibene L, Bosco D, et al. (2002) Anti-GM1 ganglioside antibodies in Parkinson's disease. Acta Neurol Scand 106: 54-57. |
| [64] | Hatano T, Saiki S, Okuzumi A, et al. (2015) Identification of novel biomarkers for Parkinson's disease by metabolomic technologies. J Neurol Neurosurg Psychiatry. |
| [65] |
Luan H, Liu LF, Meng N, et al. (2015) LC-MS-based urinary metabolite signatures in idiopathic Parkinson's disease. J Proteome Res 14: 467-478. doi: 10.1021/pr500807t
|
| [66] | Trupp M, Jonsson P, Ohrfelt A, et al. (2014) Metabolite and peptide levels in plasma and CSF differentiating healthy controls from patients with newly diagnosed Parkinson's disease. J Parkinsons Dis 4: 549-560. |
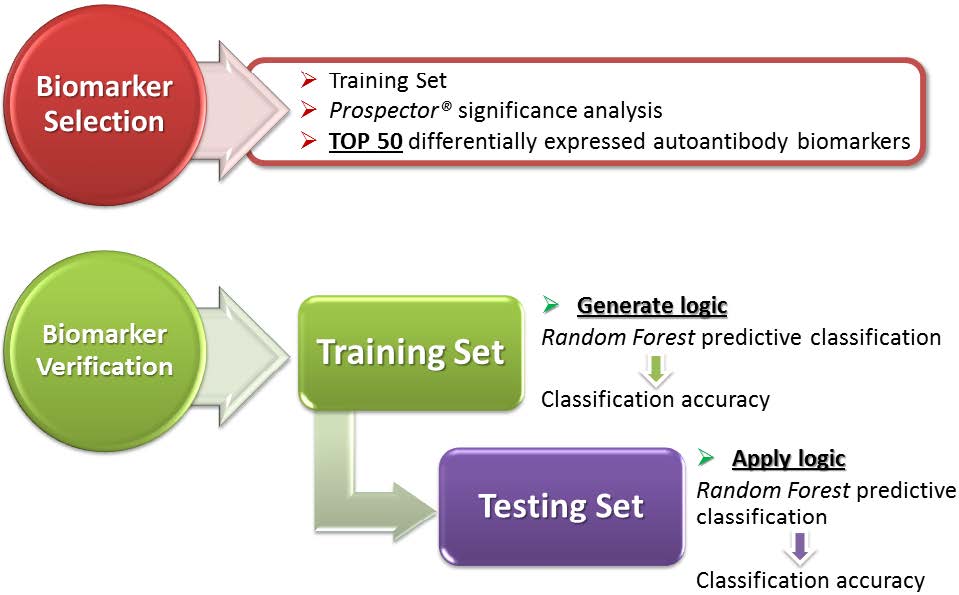
Figures(1)

Cassandra A DeMarshall, Abhirup Sarkar, Robert G Nagele. Serum Autoantibodies as Biomarkers for Parkinsons Disease: Background and Utility[J]. AIMS Medical Science, 2015, 2(4): 316-327. doi: 10.3934/medsci.2015.4.316







 DownLoad:
DownLoad: