Citation: Allison K.K. Leimer, Kenneth G. Boykin, Mark C. Andersen, Caitriana M. Steele. Applicability of functional groups as indicators of resilience and redundancy in the San Pedro Watershed, Arizona[J]. AIMS Environmental Science, 2019, 6(3): 127-146. doi: 10.3934/environsci.2019.3.127

| [1] |
Allen CR, Gunderson L, Johnson AR (2005) The use of discontinuities and functional groups to assess relative resilience in complex systems. Ecosyst 8: 958. doi: 10.1007/s10021-005-0147-x

|
| [2] |
Sundstrom SM, Allen CR, Barichievy C (2012) Species, functional groups, and thresholds in ecological resilience. Conserv Biol 26: 305–14. doi: 10.1111/j.1523-1739.2011.01822.x
|
| [3] |
Holling CS (1973) Resilience and stability of ecological systems. Annu rev ecol syst 4: 1–23. doi: 10.1146/annurev.es.04.110173.000245
|
| [4] |
Walker BH (1992) Biodiversity and ecological redundancy. Conserv biol 6: 18–23. doi: 10.1046/j.1523-1739.1992.610018.x
|
| [5] |
Walker B (1995) Conserving biological diversity through ecosystem resilience. Conserv biol 9: 747–52. doi: 10.1046/j.1523-1739.1995.09040747.x
|
| [6] |
MacArthur R (1955) Fluctuations of animal populations and a measure of community stability. Ecol 36: 533–6. doi: 10.2307/1929601
|
| [7] |
Hutchinson GE (1959) Homage to Santa Rosalia or why are there so many kinds of animals? Am Naturalist 93: 145–59. doi: 10.1086/282070
|
| [8] | May RM (1973) Stability and Complexity in Model Ecosystems. Princeton University Press Princeton, USA. |
| [9] |
McCann KS (2000) The diversity–stability debate. Nature 405: 228. doi: 10.1038/35012234
|
| [10] | Walker B, Holling CS, Carpenter SR, et al. (2004) Resilience, adaptability and transformability in social–ecological systems. Ecol Soc 9. |
| [11] |
Peterson G, Allen CR, Holling CS (1998) Ecological resilience, biodiversity, and scale. Ecosyst 1: 6–18. doi: 10.1007/s100219900002
|
| [12] | Allen CR, Holling CS (2010) Novelty, adaptive capacity, and resilience. Ecol soc 15. |
| [13] | Gunderson LH, Holling CS, Allen CR (2010) The evolution of an idea-the past, present, and future of ecological resilience. Foundations of ecological resilience 423–444. |
| [14] |
Folke C, Carpenter S, Walker B, et al. (2004) Regime shifts, resilience, and biodiversity in ecosystem management. Annu Rev Ecol Evol Syst 35: 557–81. doi: 10.1146/annurev.ecolsys.35.021103.105711
|
| [15] |
Holling CS (1992) Cross‐scale morphology, geometry, and dynamics of ecosystems. Ecol monogr 62: 447–502. doi: 10.2307/2937313
|
| [16] |
Forys EA, Allen CR (2002) Functional group change within and across scales following invasions and extinctions in the Everglades ecosystem. Ecosyst 5: 339–47. doi: 10.1007/s10021-001-0078-0
|
| [17] |
Fonseca CR, Ganade G (2001) Species functional redundancy, random extinctions and the stability of ecosystems. J Ecol 89: 118–25. doi: 10.1046/j.1365-2745.2001.00528.x
|
| [18] | Skillen JJ, Maurer BA (2008) The ecological significance of discontinuities in body-mass distributions. Discontinuities in ecosystems and other complex systems 193–218. |
| [19] |
Root RB (1967) The niche exploitation pattern of the blue‐gray gnatcatcher. Ecol monogr 37: 317–50. doi: 10.2307/1942327
|
| [20] | Kepner WG, Watts CJ, Edmonds CM, et al. (2000) A landscape approach for detecting and evaluating change in a semi-arid environment. In: Monitoring Ecological Condition in the Western United States. Dordrecht, Springer,179–195. |
| [21] | Miller SN, Kepner WG, Mehaffey MH, et al. (2002) Integrating landscape assessment and hydrologic modeling for land cover change analysis J Am Water Resources As 38: 915–29. |
| [22] |
Kepner WG, Semmens DJ, Bassett SD, et al. (2004) Scenario analysis for the San Pedro River, analyzing hydrological consequences of a future environment. Environ Monitor Assess 94: 115–27. doi: 10.1023/B:EMAS.0000016883.10110.15
|
| [23] |
Krueper D, Bart J, Rich TD (2003) Response of vegetation and breeding birds to the removal of cattle on the San Pedro River, Arizona (USA). Conserv biol 17: 607–15. doi: 10.1046/j.1523-1739.2003.01546.x
|
| [24] |
Lowry J, Ramsey RD, Thomas K, et al. (2007) Mapping moderate-scale land-cover over very large geographic areas within a collaborative framework: A case study of the Southwest Regional Gap Analysis Project (SWReGAP). Remote Sens Environ 108: 59–73. doi: 10.1016/j.rse.2006.11.008
|
| [25] |
Boykin KG, Kepner WG, Bradford DF, et al. (2013) A national approach for mapping and quantifying habitat-based biodiversity metrics across multiple spatial scales. Ecol Indic 33: 139–47. doi: 10.1016/j.ecolind.2012.11.005
|
| [26] | Boykin KG, Thompson BC, Deitner RA, et al. Predicted animal habitat distributions and species richness. Southwest Regional Gap Analysis Final Report US Geological Survey, Gap Analysis Program, Moscow, 2007. Available from: http://swregap.nmsu.edu/report/ArchiveReports/SWReGAP_Final_Report_Chapter_3_HabitatModels_Final.doc |
| [27] |
Boykin KG, Thompson BC, Propeck-Gray S (2010) Accuracy of gap analysis habitat models in predicting physical features for wildlife-habitat associations in the southwest US. Ecol Model 221: 2769–2775. doi: 10.1016/j.ecolmodel.2010.08.034
|
| [28] |
Brown JH, Nicoletto PF (1991) Spatial scaling of species composition: body masses of North American land mammals. Am Naturalist 138: 1478–512. doi: 10.1086/285297
|
| [29] | Dunning J. JB (1993) CRC Handbook of avian body masses. CRC Press, Boca Raton, Florida, USA. |
| [30] | NatureServe (2012) NatureServe Web Services. Available from: http://services.natureserve.org |
| [31] | Zammuto RM (1989) Life histories of birds: clutch size, longevity, and body mass among North American game birds. Cana J Zoo 64: 2739–49. |
| [32] |
Pineda-Munoz S, Evans AR, Alroy J (2016) The relationship between diet and body mass in terrestrial mammals. Paleobiology 42: 659–69. doi: 10.1017/pab.2016.6
|
| [33] | Pickard BR, Daniel J, Mehaffey M, et al. (2015) EnviroAtlas: A new geospatial tool to foster ecosystem services science and resource management. Syst zool 14: 45–55. |
| [34] |
Simpson GG (1964) Species density of North American recent mammals. Syst zool 13: 57–73. doi: 10.2307/2411825
|
| [35] | Steinitz C, Arias H, Bassett H, et al. (2003) Alternative futures for changing landscapes: The Upper San Pedro River Basin in Arizona and Sonora. Island Press. |
| [36] |
Isaac JL (2009) Effects of climate change on life history: implications for extinction risk in mammals. Endang species res 7: 115–23. doi: 10.3354/esr00093
|
| [37] | Gårdmark A, Enberg K, Ripa J, et al. (2003) The ecology of recovery. Annales Zoologici Fennici. Finnish Zoological and Botanical Publishing Board.131–144. |
| [38] |
Gagic V, Bartomeus I, Jonsson T, et al. (2015) Functional identity and diversity of animals predict ecosystem functioning better than species-based indices. P Roy Soc Lond B: Biol Sci 282: 20142620. doi: 10.1098/rspb.2014.2620
|
| [39] | McKerrow AJ, Tarr NM, Rubino MJ, et al. Patterns of species richness hotspots and estimates of their protection are sensitive to spatial resolution. Albright T, editor. Diversity and Distributions. Available from: http://doi.wiley.com/10.1111/ddi.12779 |
| [40] |
Luck GW, Daily GC, Ehrlich PR (2003) Population diversity and ecosystem services. Trends Ecol Evol 18: 331–6. doi: 10.1016/S0169-5347(03)00100-9
|
| [41] |
Carroll C, Noss RF, Paquet PC (2001) Carnivores as focal species for conservation planning in the Rocky Mountain region. Ecol appl 11: 961–80. doi: 10.1890/1051-0761(2001)011[0961:CAFSFC]2.0.CO;2
|
| [42] |
Sergio F, Newton I, Marchesi L (2005) Conservation: top predators and biodiversity. Nature 436: 192. doi: 10.1038/436192a
|
| [43] |
Sergio F, Caro T, Brown D, et al. (2008) Top predators as conservation tools: ecological rationale, assumptions, and efficacy. Annu rev EVOL s 39: 1–19. doi: 10.1146/annurev.ecolsys.39.110707.173545
|
| [44] |
Philpott SM, Soong O, Lowenstein JH, et al (2009) Functional richness and ecosystem services: bird predation on arthropods in tropical agroecosystems. Ecol appl 19: 1858–67. doi: 10.1890/08-1928.1
|
| [45] |
Semmens DJ, Diffendorfer JE, López-Hoffman L, et al. (2011) Accounting for the ecosystem services of migratory species: quantifying migration support and spatial subsidies. Ecol Econ 70: 2236–42. doi: 10.1016/j.ecolecon.2011.07.002
|
| [46] |
Kunz TH, Braun de Torrez E, Bauer D, et al. (2011) Ecosystem services provided by bats. Ann NY Acad Sci 1223: 1–38. doi: 10.1111/j.1749-6632.2011.06004.x
|
| [47] |
López-Hoffman L, Varady RG, Flessa KW, et al. (2010) Ecosystem services across borders: a framework for transboundary conservation policy. Frontiers in Ecol Envir 8: 84–91. doi: 10.1890/070216
|

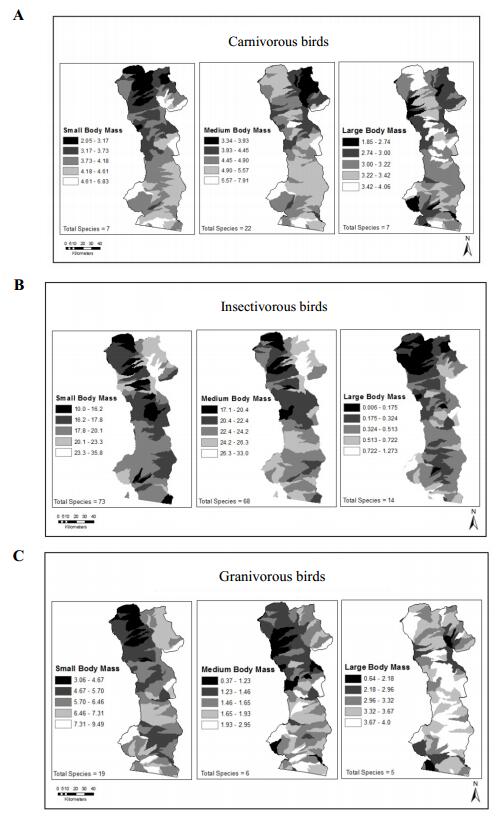
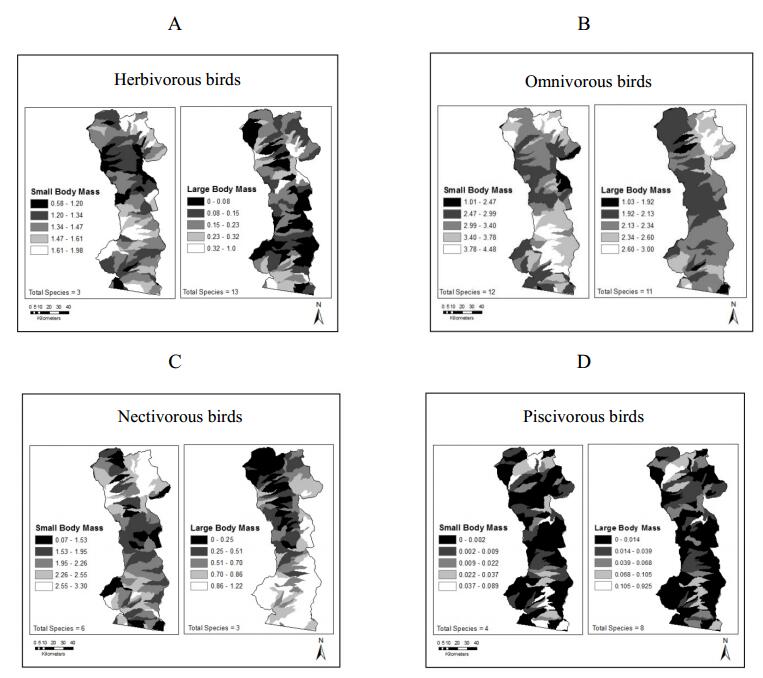


Figures(6) / Tables(3)

Allison K.K. Leimer, Kenneth G. Boykin, Mark C. Andersen, Caitriana M. Steele. Applicability of functional groups as indicators of resilience and redundancy in the San Pedro Watershed, Arizona[J]. AIMS Environmental Science, 2019, 6(3): 127-146. doi: 10.3934/environsci.2019.3.127







 DownLoad:
DownLoad: