Citation: Ranjit Kumar Upadhyay, Swati Mishra, Yueping Dong, Yasuhiro Takeuchi. Exploring the dynamics of a tritrophic food chain model with multiple gestation periods[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4660-4691. doi: 10.3934/mbe.2019234

| [1] | M. Pedraza-Garcia and L. A. Cubillos, Population dynamics of two small pelagic fish in the central-south area off Chile: delayed density-dependence and biological interaction, Env. Biol. Fish, 82 (2008), 111–122. |
| [2] | R. K. Walsh, C. Bradley, C. S. Apperson, et al., An experimental field study of delayed density dependence in natural populations of Aedes albopictus, PLoS One, 7 (2012), e35959. |
| [3] | Y. Kuang, Delay differential equations: with applications in population dynamics, Academic Press, 1993. |
| [4] | J. D. Murray, Mathematical Biology. II Spatial Models and Biomedical Applications, 3nd edition, Springer-Verlag, New York, 2003. |
| [5] | S. Gakkhar and A. Singh, Complex dynamics in a prey predator system with multiple delays, Comm. Nonlinear Sci. Numer. Simulat., 7 (2012), 914–929. |
| [6] | S. Nakaoka, Y. Saito and Y. Takeuchi, Stability, delay, and chaotic behavior in a Lotka-Volterra predator-prey system, Math. Biosci. Eng. , 3 (2006), 173–187. |
| [7] | S. Yao, L. Ding, Z. Song, et al., Two bifurcation routes to multiple chaotic coexistence in an inertial two-neural system with time delay, Nonlinear Dyn., 95 (2019), 1549–1563. |
| [8] | N. MacDonald, Biological delay systems: linear stability theory, Cambridge University Press, Uk, 1989. |
| [9] | J. M. Cushing, Integro-differential equations and delay models in population dynamics, Springer Science & Business Media, 2013. |
| [10] | Z. Song, J. Xu and B. Zhen Multitype activity coexistence in an inertial two-neuron system with multiple delays, Int. J. Bifurc. Chaos, 25 (2015), 1530040. |
| [11] | Z. Song, C. Wang and B. Zhen, Codimension-two bifurcation and multistability coexistence in an inertial two-neuron system with multiple delays, Nonlinear Dyn., 85 (2016), 2099–2113. |
| [12] | Z. Jiang, W. Zhang, J. Zhang, et al., Dynamical analysis of a phytoplankton-zooplankton system with harvesting term and Holling III functional response, Int. J. Bifur. Chaos, 28 (2018), 1850162. |
| [13] | Z. G. Song, B. Zhen and J. Xu, Species coexistence and chaotic behavior induced by multiple delays in a food chain system, Ecol. Complex, 19 (2014), 9–17. |
| [14] | G. H. Cui and X. P. Yan, Stability and bifurcation analysis on a three-species food chain system with two delays, Commun. Nonlinear Sci. Numer. Simulat., 16 (2011), 3704–3720. |
| [15] | Z. Jiang and L. Wang, Global Hopf bifurcation for a predator-prey system with three delays, Int. J. Bifur. Chaos, 27 (2017), 1750108. |
| [16] | K. Ghosh, S. Samanta, S. Biswas, et al., Stability and bifurcation analysis of an eco-epidemiological model with multiple delays, Nonlinear Stud., 23 (2016), 167–208. |
| [17] | L. Guo, Z. G. Song and J. Xu, Complex dynamics in the Leslie–Gower type of the food chain system with multiple delays, Comm. Nonlinear Sci. Numer., 19 (2014), 2850–2865. |
| [18] | R.Agrawal, D.Jana, R.K.Upadhyay, et al., Complex dynamics of sexually reproductive generalist predator and gestation delay in a food chain model: double Hopf-bifurcation to chaos, J. Appl. Math. Comput., 55 (2017), 513–547. |
| [19] | N. Bairagi and D. Jana, Dynamics and responses of a predator-prey system with competitive interference and time delay, Appl. Math. Model., 35 (2011), 3255–3267. |
| [20] | Y. Chen, J. Yu and C. Sun, Stability and Hopf bifurcation analysis in a three-level food chain system with delay, Chaos Soliton Fract., 31 (2007), 683–694. |
| [21] | Y. Dong, Y. Takeuchi and S. Nakaoka, A mathematical model of multiple delayed feedback control system of the gut microbiota-Antibiotics injection controlled by measured metagenomic data, Nonlinear Anal. Real World Appl., 43 (2018), 1–17. |
| [22] | R. K. Upadhyay and R. Agrawal, On the stability and Hopf bifurcation of a delay-induced predator-prey system with habitat complexity, Nonlinear Dyn., 83 (2017), 821–837. |
| [23] | Z. Zhang, H. Yang and H. Liu, Bifurcation analysis for a delayed food chain system with two functional responses, Electron. J. Qual. Theory Differ. Equ., 53 (2013), 1–13. |
| [24] | M. Sen M, M. Banerjee and A. Morozov, Stage-structured ratio-dependent predator-prey models revisited: When should the maturation lag result in systems? destabilization?, Ecol. Complex, 19 (2014), 23–34. |
| [25] | M. Banerjee and Y. Takeuchi, Maturation delay for the predators can enhance stable coexistence for a class of prey-predator models, J. Theor. Biol., 412 (2017), 154–171. |
| [26] | J. Wang and W. Jiang, Bifurcation and chaos of a delayed predator-prey model with dormancy of predators, Nonlinear Dyn., 69 (2012), 1541–1558. |
| [27] | D. Xiao and S. Ruan, Global analysis in a predator-prey system with nonmonotonic functional response, SIAM J. Appl. Math., 61 (2001), 1445–1472. |
| [28] | J. F. Andrews, A mathematical model for the continuous culture of microorganisms utilizing inhibitory substrates, Biotechnol. Bioeng., 10 (1968), 707–723. |
| [29] | R. Pal, D. Basu, S. Biswas, et al., Modelling of phytoplankton allelopathy with Monod–Haldane-type functional response-A mathematical study, BioSystems, 95 (2009), 243–253 |
| [30] | W. Sokol and J. A. Howell, Kinetics of phenol oxidation by washed cells, Biotechnol. Bioeng., 23 (1981), 2039–2049. |
| [31] | V. H. Edwards, Influence of high substrate concentrations on microbial kinetics, Biotechnol. Bioeng., 23 (1970), 679–712. |
| [32] | B. Boon and H. Landelout, Kinetics of nitrite oxidation by nitrobacter winogradski, Biochem J., 23 (1962), 440–447. |
| [33] | S. J. Ali, N. M. Arifin, R. K. Naji, et al., Boundedness and stability of Leslie-Gower model with Sokol-Howell functional response, Recent Advances in Math. Sci., 61 (2016), 13–26. |
| [34] | S. J. Ali, N. M. Arifin, R. K. Naji, et al., Dynamics of Leslie-Gower model with simplified Holling type IV functional response, J. Nonlinear Syst. Appl., 5 (2016), 25–33. |
| [35] | R. D. Parshad, R. K. Upadhyay, S. Mishra, et al., On the explosive instability in a three-species food chain model with modified Holling type IV functional response, Math. Methods Appl. Sci., 40 (2017), 5707–5726. |
| [36] | S. J. Ali, N. M. Arifin, R. K. Naji, et al., Analysis of ecological model with Holling type IV functional response, Int. J. Pure Appl. Math., 106 (2016), 317–331. |
| [37] | G. Huang and Y. Dong, A note on global properties for a stage structured predator-prey model with mutual interference, Adv. Differ. Equ., 2018 (2018), 308. |
| [38] | R. K. Upadhyay and R. Agrawal, Modeling the effect of mutual interference in a delay-induced predator-prey system, J. Appl. Math. Comput., 49 (2015), 13–39. |
| [39] | T. Zhang, X. Meng, Y. Song, et al., A stage-structured predator-prey SI model with disease in the prey and impulsive effects, Math. Model. Anal., 18 (2013), 505–528. |
| [40] | B. Patra, A. Maiti and G. P. Samanta, Effect of time delay on a ratio dependent food chain model, Nonlinear Anal. Model. Control, 14 (2009), 199–216. |
| [41] | N. Pal, S. Samanta, S. Biswas, et al., Stability and bifurcation analysis of a three-species food chain model with delay, Int. J. Bifur. Chaos, 25 (2015), 1550123. |
| [42] | P. J. Wangersky and W. J. Cunningham, Time lag in prey-predator population models, Ecology, 106 (1957), 136–139. |
| [43] | D. Jana, R. Agrawal and R. K. Upadhyay, Top-predator interference and gestation delay as determinants of the dynamics of a realistic model food chain, Chaos Soliton Fract., 106 (2014), 50–63. |
| [44] | J. K. Hale and S. M. Lunel, Introduction to functional differential equations, Springer Science & Business Media, 2013. |
| [45] | X. Yang, L. Chen and J. Chen, Permanence and positive periodic solution for the single-species non autonomous delay diffusive models, Comput. Math. Appl., 32 (1996), 109–116. |
| [46] | M. A. Aziz-Alaoui, Study of a Leslie-Gower-type tri-trophic population model, Chaos Soliton Fract., 14 (2002), 1275–1293. |
| [47] | Z. Song and J. Xu, Stability switches and multistability coexistence in a delay-coupled neural oscillators system, J. Theor. Biol., 313 (2012), 98–114. |
| [48] | K. McCann, A. Hastings and G. R. Huxel, Weak trophic interactions and the balance of nature, Nature, 395 (1998), 794. |
| [49] | L. Becks, F. M. Hilker, H. Malchow, et al., Experimental demonstration of chaos in a microbial food web, Nature, 435 (2005), 1226. |
| [50] | L. Becks and H. Arndt, Transitions from stable equilibria to chaos, and back, in an experimental food web, Ecology, 11 (2008), 3222–3226. |
| [51] | Y. Dong, M. Sen, M. Banerjee, et al., Delayed feedback induced complex dynamics in an Escherichia coli and Tetrahymena system, Nonlinear Dyn., 94 (2018), 1447–1466. |
| [52] | R. K. Upadhyay and R. K. Naji, Dynamics of a three species food chain model with Crowley-Martin type functional response, Chaos Soliton Fract., 42 (2009), 1337–1346. |
| [53] | R. K. Upadhyay and S. N. Raw, Complex dynamics of a three species food-chain model with Holling type IV functional response, Nonlinear Anal. Model Control, 16 (2011), 353–374. |
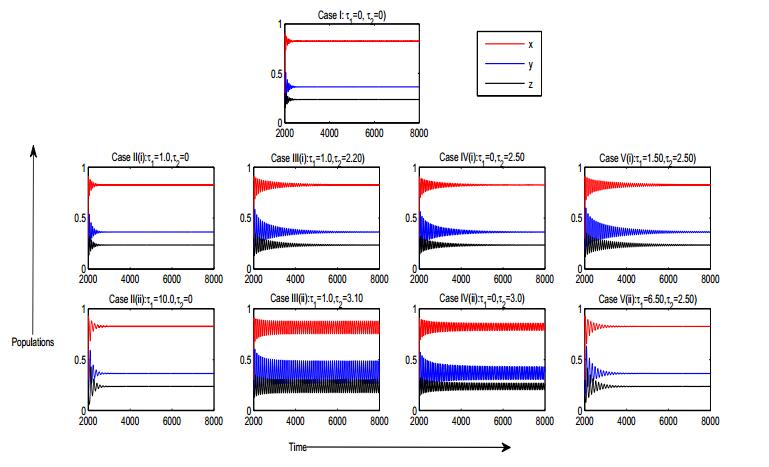
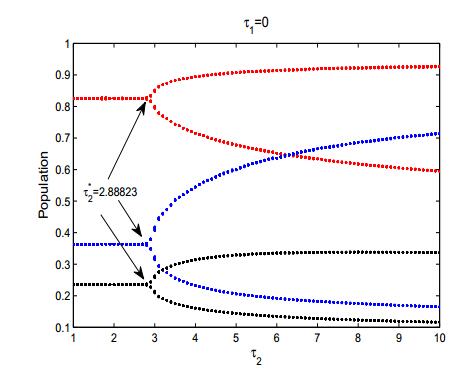
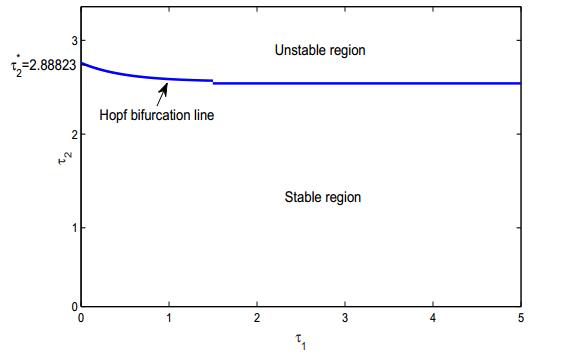
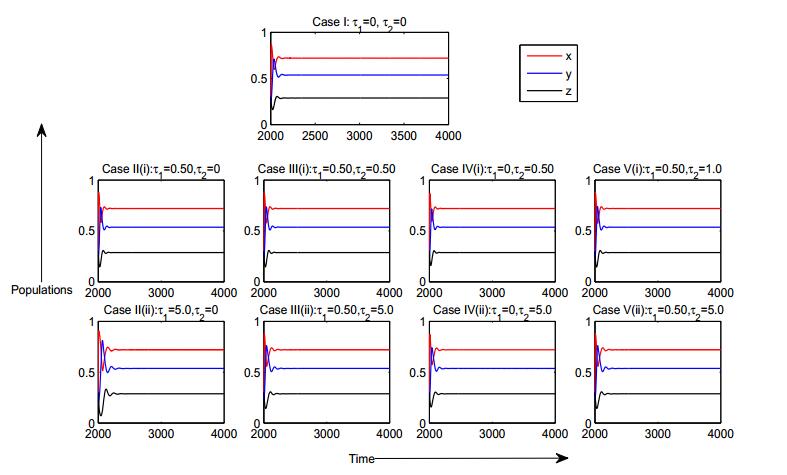
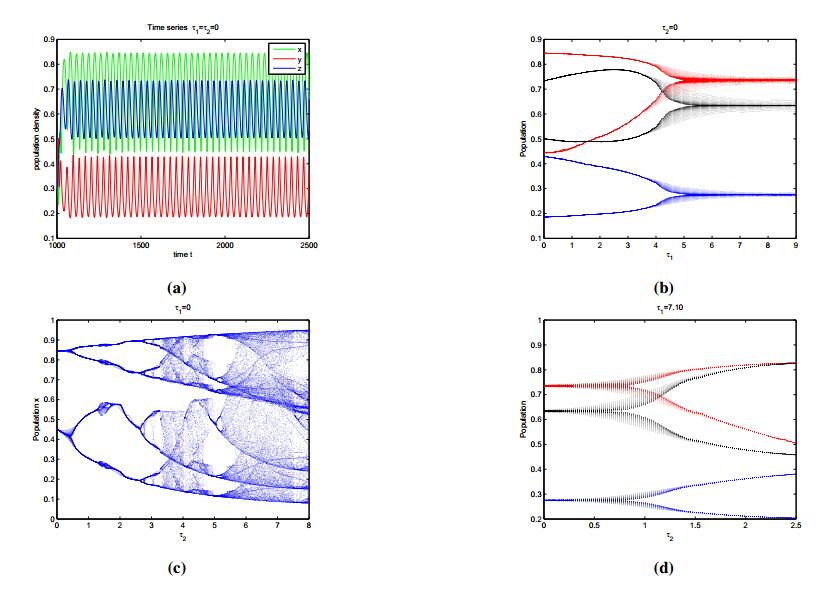
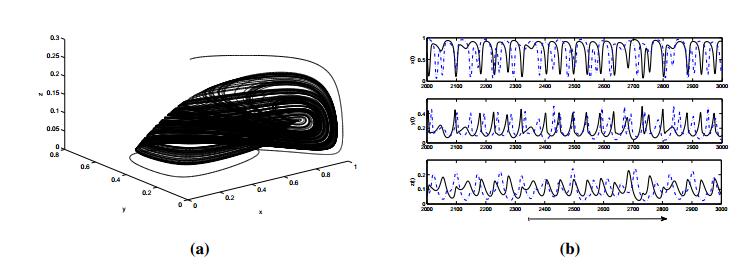
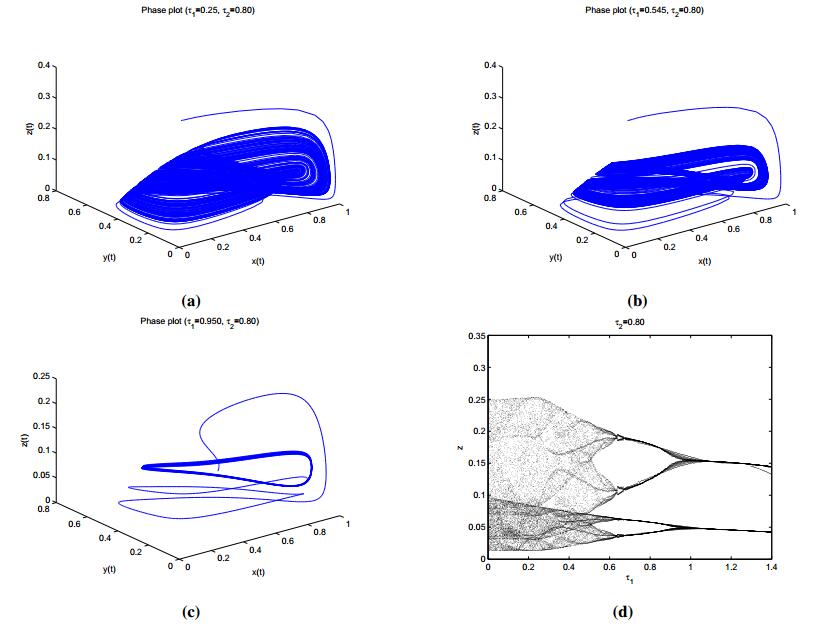
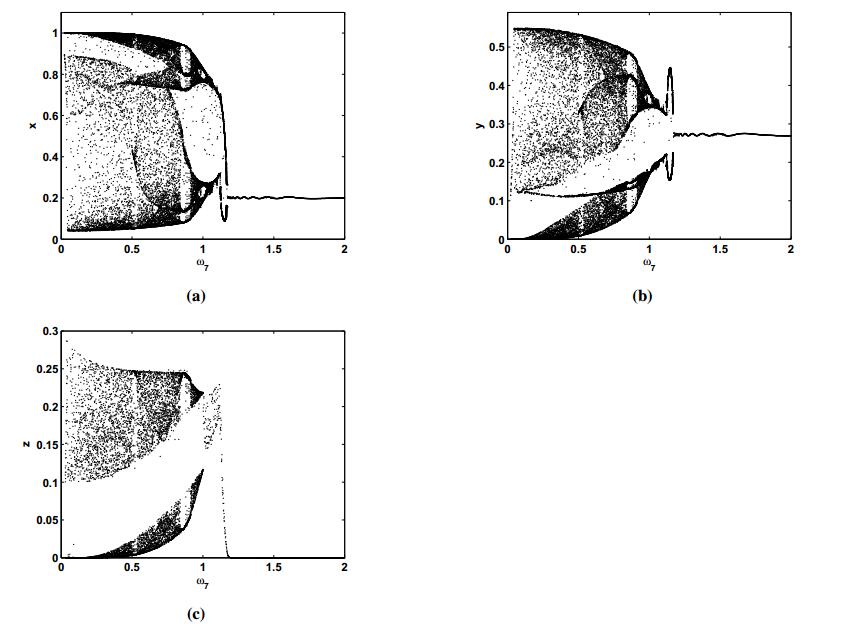
Figures(8) / Tables(2)

Ranjit Kumar Upadhyay, Swati Mishra, Yueping Dong, Yasuhiro Takeuchi. Exploring the dynamics of a tritrophic food chain model with multiple gestation periods[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4660-4691. doi: 10.3934/mbe.2019234







 DownLoad:
DownLoad: