Citation: Jeremy Bingham, Pietro Landi, Cang Hui. Prejudice, privilege, and power: Conflicts and cooperation between recognizable groups[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4092-4106. doi: 10.3934/mbe.2019203

| [1] | T. Clutton-Brock, Cooperation between non-kin in animal societies, Nature, 462 (2009), 51–57. |
| [2] | R. L. Trivers, The evolution of reciprocal altruism, Q. Rev. Biol., 46 (1971), 35–57. |
| [3] | C. Darwin, On the origin of species by means of natural selection, or preservation of favoured races in the struggle for life. John Murray, London, 1859. |
| [4] | C. Hauert, M. Holmes and M. Doebeli, Evolutionary games and population dynamics: maintenance of cooperation in public goods games, Proc. R. Soc. B, 273 (2006), 2565–2570. |
| [5] | W. D. Hamilton, The genetical ecolution of social behavior I and II, J. Theor. Biol., 7 (1964), 1–52. |
| [6] | R. Axelrod, The Evolution of Cooperation, Basic Books, Inc., New York, 1981. |
| [7] | R. Boyd and P. J. Richerson, The evolution of indirect reciprocity, Social Networks, 11 (1989), 213 – 236. |
| [8] | M. A. Nowak and K. Sigmund, Evolution of indirect reciprocity, Nature, 437 (2005), 1291–1298. |
| [9] | A. Gardner and S. A. West, Greenbeards, Evolution, 64 (2009), 25–38. |
| [10] | S. D. Carroll, Evolutionary games and population dynamics, Q. Rev. Biol., 74 (1999), 347–347. |
| [11] | R. Boyd and P. J. Richerson, Culture and the evolution of human cooperation, Philos. Trans. R. Soc. B, 364 (2009), 3281–3288. |
| [12] | L. Liu, S. Wang, X. Chen, et al., Evolutionary dynamics in the public goods games with switching between punishment and exclusion, Chaos, 28 (2018), 103105. |
| [13] | X. Chen, T. Sasaki and M. Perc, Evolution of public cooperation in a monitored society with implicated punishment and within-group enforcement, Sci. Rep., 5 (2011). |
| [14] | M. Perc, J. Gomez-Gardenes, A. Szolnoki, et al., Evolutionary dymanics of group interactions on structured populations: a review, J. R. Soc. Interface, 10 (2013), 20120997. |
| [15] | A. Szolnoki and M. Perc, Second-order free-riding on antisocial punishment restores the effectiveness of prosocial punishment, Phys. Rev. X, 7 (2017), 198192. |
| [16] | K. Mahmoodi, B. J. West and P. Grigolini, Self-organizing Complex Networks: individual versus global rules, Front. Physiol., 8 (2017), 478. |
| [17] | M. Perc, J. J. Jordan, D. G. Rand, et al., Statistical physics of human cooperation Phys. Rep., 687 (2017), 1–51. |
| [18] | Z. Wang, L. Wang, A. Szolnoki, et al., Evolutionary games on multilayer networks: a colloquium,Eur. Phys. J. B, 88 (2015), 124. |
| [19] | M. A. Nowak, Five rules for the evolution of cooperation., Science, 314 (2006), 1560–1563. |
| [20] | K. M. Page and M. A. Nowak, Unifying evolutionary dynamics, J. Theor. Biol., 219 (2002), 93–98. |
| [21] | M. van Baalend and D. A. Rand The unit of selection in viscous populations and the evolution of altruism, J. Theor. Biol., 193 (1998), 631–648. |
| [22] | F. Zhang and C. Hui, Eco-evolutionary feedback and the invasion of cooperation in prisoner's dilemma games, PLoS One, 6 (2011), e27523. |
| [23] | F. Zhang, Y. Tao, Z. Li, et al., The evolution of cooperation on fragmented landscapes: the spatial hamilton rule, Evol. Ecol. Res., 12 (2010), 23–33. |
| [24] | P. Richerson, A .V. Bell, and R. Baldini et al., Cultural group selection plays an essential role in explaining human cooperation: A sketch of the evidence, Behav. Brain Sci., 29 (2016), 30. |
| [25] | B. C. Eaton, M. Eswaran and R. J. Oxoby, 'Us' and 'Them': The origin of identity, and its economic implications, Can. J. Econ., 44 (2011), 719–748. |
| [26] | R. M. Kramer and M. B. Brewer, Effects of group identity on resource use in a simulated commons dilemma, J. Pers. Soc. Psychol., 46 (1984), 1044–1057. |
| [27] | M. B. Brewer and R. M. Kramer, Choice behavior in social dilemmas: Effects of social identity, group size, and decision framing, J. Pers. Soc. Psychol., 50 (1986), 543–549. |
| [28] | G. Hardin, The trajedy of the commons, Science, 162 (1968), 1243–1248. |
| [29] | D. H. Cole and P. Z. Grossman, Institutions matter! why the herder problem is not a prisoner's dilemma, Theo. Dec., 69 (2008), 219–231. |
| [30] | R. H. McAdams, Beyond the prisoners' dilemma: Coordination, game theory, and law, South. Calif. Law Rev., 82 (2009), 209. |
| [31] | B. Skyrms, The Stag Hunt and the Evolution of Social Structure, Cambridge University Press, 2003. |
| [32] | J.-J. Rousseau, R. D. Masters and C. Kelly, Discourse on the origins of inequality; polemics; and, political economy. Hanover, NH, Published for Dartmouth College by University Press of New England, 1992. |
| [33] | T. Antal, H. Ohtsuki, J. Wakeley, et al., Evolution of cooperation by phenotypic similarity, Proc. Natl. Acad. Sci. USA, 106 (2009), 8597–8600. |
| [34] | F. Fu, C. E. Tarnita, N. A. Christakis, et al., Evolution of in-group favoritism, Sci. Rep., 2 (2012), 460. |
| [35] | F. Jansson, What games support the evolution of an ingroup bias?, J. Theor. Biol., 373 (2015), 100–110. |
| [36] | W. A. Darity Jr, P. L. Mason and J. B. Stewart, The economics of identity: The origin and persistence of racial identity norms, J. Econ. Behav. Organ., 60 (2006), 283–305. |
| [37] | F. M. Kai and A. Konrad, Evolutionarily stable in-group favoritism and out-group spite in intergroup conflict, J. Theor. Biol., 306 (2012), 61–67. |
| [38] | N. Masuda, Ingroup favoritism and intergroup cooperation under indirect reciprocity based on group reputation, J. Theor. Biol., 311 (2012), 8–18. |
| [39] | K. Pattni, M. Broom and J. Rychtář, Evolutionary dynamics and the evolution of multiplayer cooperation in a subdivided population, J. Theor. Biol., 429 (2017), 105–115. |
| [40] | R. L. Riolo, M. D. Cohen and R. Axelrod, Evolution of cooperation without reciprocity, Nature,414 (2001), 441–443. |
| [41] | J. P. Bruner, Diversity and cooperation, Ph.D thesis, UC Irvine, 2014. |
| [42] | S. T. Powers, D. J. Taylor and J. J. Bryson, Punishment can promote defection in group-structured populations, J. Theor. Biol., 311 (2012), 107–116. |
| [43] | C. Hui, P. Landi, H. O. Minoarivelo, et al., Ecological and Evolutionary Modelling, Springer International Publishing, 2018. |
| [44] | N. Alexander, Affirmative action and the perpetuation of racial identities in post-apartheid South Africa, Edited version of a Lecture originally delivered at the East London Campus, University of Fort Hare, 2006. |
| [45] | K. Durrheim, M. Xoliswa and L. Brown, Race trouble: race, identity and inequality in post- apartheid South Africa, Lexington Books, 2011. |
| [46] | R. A. Wilson, The politics of truth and reconciliation in South Africa: Legitimizing the post- apartheid state, Cambridge University Press, 2001. |
| [47] | N. Alexander, Language policy and national unity in South Africa/Azania, Buchu Books, Cape Town, 1989. |
| [48] | N. W. Thiong'o, Decolonizing the mind: the politics of language in African literature, James Currey, London, 1986. |
| [49] | P. Landi and F. Dercole, The social diversification of fashion, J. Math. Soc., 40 (2016), 185–205. |
| [50] | M. Hartshorn, A. Kaznatcheev and T. Schultz, The evolutionary dominance of ethnocentric cooperation, J. Artif. Soc. S., 16 (2013), 7. |
| [51] | R. Dawkins, The Selfish Gene, Oxford University Press, Oxford, UK, 1976. |
| [52] | V. A. A. Jansen and M. van Baalen, Altruism through beard chromodynamics, Nature, 440 (2006), 663. |
| [53] | A. Traulsen and M. A. Nowak, Chromodynamics of cooperation in finite populations, PLoS One,2 (2007), e270. |
| [54] | C. Hui, H. O. Minoarivelo and P. Landi, Modelling coevolution in ecological networks with adaptive dynamics, Math. Method. Appl. Sci., 41 (2018), 8407–8422. |
| [55] | M. Perc and A. Szolnoki Coevolutionary games - a mini review, BioSystems, 99 (2010), 109–125. |
| [56] | V. Capraro and M. Perc, Grand challenges in social physics: in pursuit of moral behavior, Front. Phys., 6 (2018), 107. |
| [57] | K. Mahmoodi, P. Grigolini and B. J. West, On social sensitivity to either zealot or independent minorities, Chaos, Soliton. Fract., 110 (2018), 185–190. |
| [58] | P. Landi and C. Piccardi, Community analysis in directed networks: in-, out- and pseudocommunities, Phys. Rev. E, 89 (2014), 012814. |
| [59] | K. Mahmoodi and P. Grigolini Evolutionary game theory and criticality J. Phys. A- Math. Theor., 50 (2017), 015101. |
| [60] | M.A.NowakandR.M.May, Evolutionarygamesandspatialchaos, Nature, 359(1992), 826–829.v 61. J. J. Horton, D. G. Rand and R. J. Zeckhauser, The online laboratory: conducting experiments in a real labor market. Exp. Econ., 14 (2011), 399–425. |
| [61] | 62. A. Ofra and D. G. Rand, Economic games on the internet: the effect of $1 stakes, PloS One, 7 (2012), e314 |
| [62] | 63. D. G. Rand, The promise of Mechanical Turk: how online labor markets can help theorists run behavioural experiments, J. Theor. Biol., 299 (2012), 172–179. |
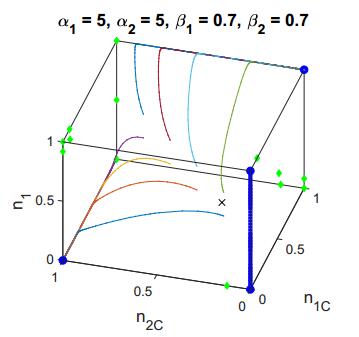
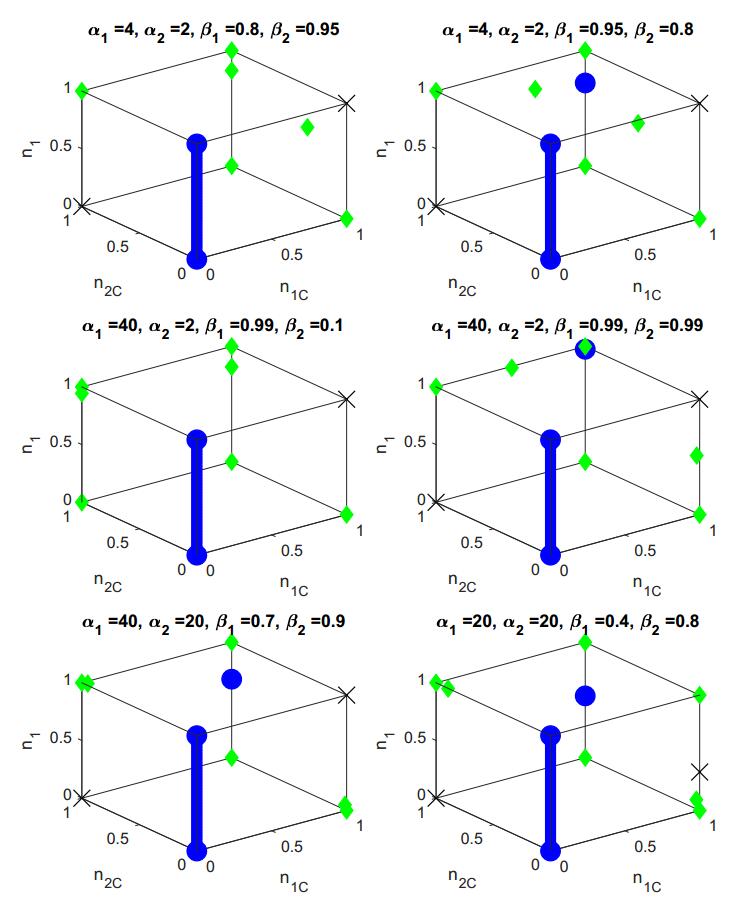
Figures(2) / Tables(3)

Jeremy Bingham, Pietro Landi, Cang Hui. Prejudice, privilege, and power: Conflicts and cooperation between recognizable groups[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4092-4106. doi: 10.3934/mbe.2019203







 DownLoad:
DownLoad: