D-pinitol is a natural compound related to the important family of inositols. It can be found and isolated from many plants, being the active component of ayurvedic remedies such as Talisa patra (Abies webbiana, A. pindrow) or antidiabetic as Bougainvillea (Bougainvillea spectabilis). Although many synthetic and semi-synthetic methods have been reported for D-pinitol and its derivatives, through chemical and biochemical transformations, Ceratonia siliqua L. (Carob), a Mediterranean tree now in decline, known because of its environmental advantages, is the only raw material from which D-pinitol can be isolated in quantities enough for a viable commercial exploitation. Fortunately, the pharmacological interest in this compound has risen enormously in the last years owing to their established multifunctional properties through a variety of signalling pathways: ⅰ) anti-cancer, through inhibition of TNF-ᾳ and suppression of NF-ⱪB pathway; ⅱ) insulinomimetic and metabolic regulator in type 2 diabetes mellitus, via a post-receptor pathway of insulin action; ⅲ) antioxidant; ⅳ) hepatoprotective; ⅴ) immuno-modulator, balancing Th1/Th2 cytokines; ⅵ) osteoporosis preventive, through p38/JNK and NF-ⱪB pathways; ⅶ) anti-aging, via reduction of the insulin/IGF-1 signaling (IIS) pathway; ⅷ) improver of creatine retention; ⅸ) preventive and ameliorative of Alzheimer's disease through selective g-secretase modulation.
Thus, the present review compress the literature reported to date in relation to the health-promoting effects and metabolic pathways of this naturally occurring super-food ingredient and its derivatives, providing an extensive guide for a future utilization of all of its potentialities, aiming a positive impact in the promotion and recovery of carob crops.
Citation: JoséIgnacio López-Sánchez, Diego A. Moreno, Cristina García-Viguer. D-pinitol, a highly valuable product from carob pods: Health-promoting effects and metabolic pathways of this natural super-food ingredient and its derivatives[J]. AIMS Agriculture and Food, 2018, 3(1): 41-63. doi: 10.3934/agrfood.2018.1.41
D-pinitol is a natural compound related to the important family of inositols. It can be found and isolated from many plants, being the active component of ayurvedic remedies such as Talisa patra (Abies webbiana, A. pindrow) or antidiabetic as Bougainvillea (Bougainvillea spectabilis). Although many synthetic and semi-synthetic methods have been reported for D-pinitol and its derivatives, through chemical and biochemical transformations, Ceratonia siliqua L. (Carob), a Mediterranean tree now in decline, known because of its environmental advantages, is the only raw material from which D-pinitol can be isolated in quantities enough for a viable commercial exploitation. Fortunately, the pharmacological interest in this compound has risen enormously in the last years owing to their established multifunctional properties through a variety of signalling pathways: ⅰ) anti-cancer, through inhibition of TNF-ᾳ and suppression of NF-ⱪB pathway; ⅱ) insulinomimetic and metabolic regulator in type 2 diabetes mellitus, via a post-receptor pathway of insulin action; ⅲ) antioxidant; ⅳ) hepatoprotective; ⅴ) immuno-modulator, balancing Th1/Th2 cytokines; ⅵ) osteoporosis preventive, through p38/JNK and NF-ⱪB pathways; ⅶ) anti-aging, via reduction of the insulin/IGF-1 signaling (IIS) pathway; ⅷ) improver of creatine retention; ⅸ) preventive and ameliorative of Alzheimer's disease through selective g-secretase modulation.
Thus, the present review compress the literature reported to date in relation to the health-promoting effects and metabolic pathways of this naturally occurring super-food ingredient and its derivatives, providing an extensive guide for a future utilization of all of its potentialities, aiming a positive impact in the promotion and recovery of carob crops.

| [1] |
Nasar-Abbas SM, E-Huma Z, Vu T, et al. (2016) Carob Kibble: A Bioactive-Rich Food Ingredient. Compr Rev Food Sci Food Saf 15: 63-72. doi: 10.1111/1541-4337.12177

|
| [2] |
Bhat KA, Shah BA, Gupta KK, et al. (2009) Semi-synthetic analogs of pinitol as potential inhibitors of TNF-alpha cytokine expression in human neutrophils. Bioorg Med Chem Lett 19: 1939-1943. doi: 10.1016/j.bmcl.2009.02.050
|
| [3] |
Anderson AB, MacDonald DL, Fischer HOL (1952) The structure of pinitol. J Am Chem Soc 74: 1479-1480. doi: 10.1021/ja01126a036
|
| [4] |
Posternak T (1936) Recherches dans la série des cyclites Ⅲ. Sur la configuration des inosites actives. Helv Chim Acta 19: 1007-1010. doi: 10.1002/hlca.193601901132
|
| [5] |
Catelani G, D'Andrea F, Griselli A, et al. (2008) A new stereoselective approach to a selectively protected derivative of D-pinitol and its evaluation as alpha-L-rhamnopyranose mimetic. Tetrahedron Lett 49: 4534-4536. doi: 10.1016/j.tetlet.2008.05.040
|
| [6] |
Dowd M, Stevens E (2002) The crystal structures of D-Pinitol and 1-Quebrachitol by low-temperature X-ray diffraction. J Carbohydr Chem 21: 373-383. doi: 10.1081/CAR-120014901
|
| [7] | Dowd MK, Stevens ED, Experimental Crystal Structure Determination. CCDC 172582,2014. Available from: https://dx.doi.org/10.5517/cc5sl5p. |
| [8] |
Raya-Gonzalez D, Pamatz-Bolanos T, Rio-Torres R, et al. (2008) D-(+)-Pinitol, a component of the heartwood of Enterolobium cyclocarpum (Jacq.) Griseb. Z Naturforsch C: J Biosci 63: 922-924. doi: 10.1515/znc-2008-11-1225
|
| [9] | Anderson I, (1972) The cyclitols, In: Pigman W, Horton D. Eds. The Carbohydrates, 2nd ed., New York and London: Academic Press, Inc., Vol. 1A Chemistry and Biochemisty. |
| [10] |
Calle J, Rivera A, Josephnathan P (1987) Pinitol from leaves of Gliricidia sepium. Planta Med 53: 303. doi: 10.1055/s-2006-962717
|
| [11] | Poongothai G, Sripathi SK (2013) A review on insulinomimetic pinitol from plants. Int J Pharm Bio Sci 4: 992-1009. |
| [12] |
Al-Suod H, Lior M, Ratiu IA, et al. (2016) A window on cyclitols: Characterization and analytics of inositols. Phytochem Lett 20: 507-519. doi: 10.1016/j.phytol.2016.12.009
|
| [13] |
Labed A, Ferhat M, Labed-Zouad I, et al. (2016) Compounds from the pods of Astragalus armatus with antioxidant, anticholinesterase, antibacterial and phagocytic activities. Pharm Biol 54: 3026-3032. doi: 10.1080/13880209.2016.1200632
|
| [14] | Lahuta LB, Ciak M, Rybinski W, et al. (2017) Diversity of the composition and content of soluble carbohydrates in seeds of the genus Vicia (Leguminosae). Genet Resour Crop Evol 2017: 1-14. |
| [15] |
Phillips DV, Dougherty DE, Smith AE (1982) Cyclitols in soybean. J Agric Food Chem 30: 456-458. doi: 10.1021/jf00111a011
|
| [16] |
Adinarayana D, Ramachandraiah P (1985) C-Glycosylphenolics from Rhynchosia suaveolens. J Nat Prod 48: 156-157. doi: 10.1021/np50037a042
|
| [17] |
Tetik N, Turhan I, Oziyci HR, et al. (2011) Determination of D-pinitol in carob syrup. Int J Food Sci Nutr 62: 672-676. doi: 10.3109/09637486.2011.560564
|
| [18] |
Sharma N, Verma MK, Gupta DK, et al. (2016) Isolation and quantification of pinitol in Argyrolobium roseum plant, by 1H-NMR. J Saudi Chem Soc 20: 81-87. doi: 10.1016/j.jscs.2014.07.002
|
| [19] | Pérez-Pastor A, Soares-Neto JP, de la Rosa JM, et al. (2016) Carbon footprint assessment in carob tree plantations. Vida Rural April: 52-60. |
| [20] |
Baumgartner S, Gennerritzmann R, Haas J, et al. (1986) Isolation and identification of cyclitols in carob pods (Ceratonia siliqua L.). J Agric Food Chem 34: 827-829. doi: 10.1021/jf00071a015
|
| [21] | Oziyci HR, Turhan I, Tetik N (2015) Concentration of D-pinitol in carob extract by using multi-stage enrichment processes. GIDA 40: 125-131. |
| [22] | Turhan I (2011) Optimization of extraction of D-pinitol and phenolics from cultivated and wild types of carob pods using response surface methodology. Int J Food Eng 7: 639-646. |
| [23] |
Turhan I (2014) Relationship between sugar profile and D-pinitol content of pods of wild and cultivated types of carob bean (Ceratonia siiqua L.). Int J Food Prop 17: 363-370. doi: 10.1080/10942912.2011.631255
|
| [24] |
Lin TH, Tan TW, Tsai TH, et al. (2013) D-pinitol inhibits prostate cancer metastasis through inhibition of aVb3 integrin by modulating FAK, c-Src and NF-kB pathways. Int J Mol Sci 14: 9790-9802. doi: 10.3390/ijms14059790
|
| [25] |
Streeter JG (1980) Carbohydrates in soybean nodules: Ⅱ. Distribution of compounds in seedlings during the onset of nitrogen fixation. Plant Physiol 66: 471-476. doi: 10.1104/pp.66.3.471
|
| [26] |
Hudlicky T, Price JD, Rulin F, et al. (1990) Efficient and enantiodivergent synthesis of (+)-and (-)-pinitol. J Am Chem Soc 112: 9439-9440. doi: 10.1021/ja00181a081
|
| [27] |
Hudlicky T, Rulin F, Tsunoda T, et al. (1991) Biocatalysis as a rational approach to enantiodivergent synthesis of highly oxygenated compounds: (+)-and (-)-Pinitol and Other Cyclitols. Isr J Chem 31: 229-238. doi: 10.1002/ijch.199100027
|
| [28] |
Hudlicky T, Mandel M, Rouden J, et al. (1994) Microbial Oxidation of Aromatics in Enantiocontrolled Synthesis. Part 1. Expedient and General Asymmetric Synthesis of lnositols and Carbohydrates via an Unusual Oxidation of a Polarized Diene with Potassium Permanganate. J Chem Soc Perkin Trans 1 1: 1553-1567. doi: 10.1039/p19940001553
|
| [29] |
Ley SV, Sternfeld F (1989) Microbial oxidation in synthesis: Preparation of (+)-and (-)-pinitol from benzene. Tetrahedron 45: 3463-3476. doi: 10.1016/S0040-4020(01)81025-5
|
| [30] |
Ley SV, Sternfeld F, Taylor S (1987) Microbial oxidation in synthesis: A six step preparation of (+)-Pinitol from benzene. Tetrahedron Lett 28: 225-226. doi: 10.1016/S0040-4039(00)95692-2
|
| [31] |
Aceña JL, Arjona O, Plumet J (1996) Total synthesis of (+)-pinitol. Tetrahedron: Asymmetry 7: 3535-3544. doi: 10.1016/S0957-4166(96)00461-2
|
| [32] |
Sethi G, Ahn KS, Sung B, et al. (2008) Pinitol targets nuclear factor-kB activation pathway leading to inhibition of gene products associated with proliferation, apoptosis, invasion, and angiogenesis. Mol Cancer Ther 7: 1604-1614. doi: 10.1158/1535-7163.MCT-07-2424
|
| [33] |
Gao Y, Zhang M, Wu T, et al. (2015) Effects of D-pinitol on insulin resistance through the PI3K/Akt signaling pathway in type 2 diabetes mellitus rats. J Agric Food Chem 63: 6019-6026. doi: 10.1021/acs.jafc.5b01238
|
| [34] |
Liu SC, Chuang SM, Tang CH (2012) D-pinitol inhibits RANKL-induced osteoclasteogenesis. Int Immunopharmacol 12: 494-500. doi: 10.1016/j.intimp.2012.01.002
|
| [35] |
Zhou Y, Park CM, Cho CW, et al. (2008) Protective effect of pinitol against D-galactosamine-induced hepatotoxicity in rats fed on a high-fat diet. Biosci Biotechnol Biochem 72: 1657-1666. doi: 10.1271/bbb.70473
|
| [36] | Shin YC, Jeon JY (2004) The physiological activities of pinitol isolated from soybean. Food Ind Nutr 30: 2680-2688. |
| [37] |
Choi MS, Lee MK, Jung UJ, et al. (2009) Metabolic response of soy pinitol on lipid-lowering, antioxidant and hepatoprotective action in hamsters fed-high fat and high cholesterol diet. Mol Nutr Food Res 53: 751-759. doi: 10.1002/mnfr.200800241
|
| [38] |
Rengarajan T, Nandakumar N, Rajendran P, et al. (2014) D-pinitol promotes apoptosis in MCF-7 cells via induction of p53 and Bax and inhibition of Bcl-2 and NF-kB. Asian Pac J Cancer Prev 15: 1757-1762. doi: 10.7314/APJCP.2014.15.4.1757
|
| [39] |
Rengarajan T, Nandakumar N, Rajendran P, et al. (2015) D-pinitol mitigates tumor growth by modulating interleukins and hormones and induces apoptosis in rat breast carcinogenesis through inhibition of NF-kB. J Physiol Biochem 71: 191-204. doi: 10.1007/s13105-015-0397-9
|
| [40] | Fentiman IS (2001) Fixed and modifiable risk factors for breast cancer. Int J Clin Pract 55: 527-530. |
| [41] |
Parkin DM, Bray F, Ferlay J, et al. (2001) Estimating the world cancer burden: Globocan 2000. Int J Cancer 94: 153-156. doi: 10.1002/ijc.1440
|
| [42] |
Mundy GR (2002) Metastasis: Metastasis to bone: Causes, consequences and therapeutic opportunities. Nat Rev Cancer 2: 584-593. doi: 10.1038/nrc867
|
| [43] |
Bryant RJ, Hamdy FC (2008) Screening for prostate cancer: An update. Eur Urol 53: 37-44. doi: 10.1016/j.eururo.2007.08.034
|
| [44] |
Ernst DS, Hanson J, Venner PM (1991) Analysis of prognostic factors in men with metastatic prostate cancer. Uro-Oncology Group of Northern Alberta. J Urol 146: 372-376. doi: 10.1016/S0022-5347(17)37797-2
|
| [45] |
Jayasooriya R, Kang CK, Park SR, et al. (2015) Pinitol suppresses tumor necrosis factor-a-induced invasion of prostate cancer LNCaP cells by inhibiting nuclear factor-kB-Mediated matrix metalloproteinase-9 expression. Trop J Pharm Res 14: 1357-1364. doi: 10.4314/tjpr.v14i8.6
|
| [46] |
Ayala GE, Dai H, Ittmann M, et al. (2004) Growth and survival mechanisms associated with perineural invasion in prostate cancer. Cancer Res 64: 6082-6090. doi: 10.1158/0008-5472.CAN-04-0838
|
| [47] |
Nakamachi H, Suzuki H, Akakura K, et al. (2002) Clinical significance of pulmonary metastases in stage D2 prostate cancer patients. Prostate Cancer Prostatic Dis 5: 159-163. doi: 10.1038/sj.pcan.4500573
|
| [48] |
Pezzuto JM (1997) Plant-derived anticancer agents. Biochem Pharmacol 53: 121-133. doi: 10.1016/S0006-2952(96)00654-5
|
| [49] |
Conney AH, Lou YR, Xie JG, et al. (1997) Some perspectives on dietary inhibition of carcinogenesis: Studies with curcumin and tea. Proc Soc Exp Biol Med 216: 243-245. doi: 10.3181/00379727-216-44173
|
| [50] |
Park EJ, Pezzuto JM (2002) Botanicals in cancer chemoprevention. Cancer Metastasis Rev 21: 231-255. doi: 10.1023/A:1021254725842
|
| [51] | Available from: a) https://www.cancer.gov/about-cancer/treatment/cam/patient/suns-soup-pdq. b) https://www.cancer.gov/about-cancer/treatment/cam/patient/prostate-supplements-pdq#section/_95. |
| [52] |
Rengarajan T, Nandakumar N, Balasubramanian MP (2013) D-pinitol prevents rat breast carcinogenesis induced by 7, 12-dimethylbenz (a) anthracene through inhibition of Bcl-2 and induction of p53, caspase-3 proteins and modulation of hepatic biotransformation enzymes and antioxidants. Biomed Prev Nutr 3: 31-41. doi: 10.1016/j.bionut.2012.07.001
|
| [53] | Kim YS, Park JS, Kim MJ, et al. (2014) Inhibitory effect of D-pinitol on both growth and recurrence of breast tumor from MDA-MB-231 Cancer Cells. Korean J Pharmacogn 45: 174-180. |
| [54] |
Rengarajan T, Nandakumar N (2012) Protective efficacy of dietary D-pinitol on hepatic and renal tissues during experimental breast cancer in rats challenged with 7, 12-Dimethylbenz (a) anthracene: A biochemical approach. Biomed Aging Pathol 2: 85-93. doi: 10.1016/j.biomag.2012.07.008
|
| [55] |
Rengarajan T, Rajendran P, Nandakumar N, et al. (2014) Free radical scavenging and antioxidant activity of D-pinitol against 7, 12-Dimethylbenz (a) anthracene induced breast cancer in sprague dawley rats. Asian Pac J Trop Dis 4: 384-390. doi: 10.1016/S2222-1808(14)60592-2
|
| [56] | Rengarajan T, Nandakumar N, Balasubramanian MP (2012) D-pinitol attenuates 7, 12-dimethylbenz (a) anthracene induced hazards through modulating protein bound carbohydrates, adenosine triphosphatases and lysosomal enzymes during experimental mammary carcinogenesis. J Exp Ther Oncol 10: 39-49. |
| [57] | Song SG, Park JS, Kim YS (2015) Use of pinitol and D-chiro inositol in cancer treatment and prevention of cancer relapse. Korean patent KR 20150088589 (A). |
| [58] |
Kumar A, Takada Y, Boriek AM, et al. (2004) Nuclear factor-kB: Its role in health and disease. J Mol Med 82: 434-448. doi: 10.1007/s00109-004-0555-y
|
| [59] |
Chaturvedi MM, Sung B, Yadav VR, et al. (2011) NF-kB addiction and its role in cancer: 'One size does not fit all'. Oncogene 30: 161-1630. doi: 10.1038/onc.2010.566
|
| [60] |
Kong D, Li Y, Wang Z, et al. (2007) Inhibition of angiogenesis and invasion by 3, 3'-diindolylmethane is mediated by the nuclear factor-kB downstream target genes MMP-9 and uPA that regulated bioavailability of vascular endothelial growth factor in prostate cancer. Cancer Res 67: 3310-3319. doi: 10.1158/0008-5472.CAN-06-4277
|
| [61] |
Lechertier T, Hodivala-Dilke K (2012) Focal adhesion kinase and tumour angiogenesis. J Pathol 226: 404-412. doi: 10.1002/path.3018
|
| [62] |
Hwangbo C, Kim J, Lee JJ, et al. (2010) Activation of the integrin effector kinase focal adhesion kinase in cancer cells is regulated by crosstalk between protein kinase Calpha and the PDZ adapter protein mda-9/Syntenin. Cancer Res 70: 1645-1655. doi: 10.1158/0008-5472.CAN-09-2447
|
| [63] |
Boukerche H, Su ZZ, Prévot C, et al. (2008) Mda-9/Syntenin promotes metastasis in human melanoma cells by activating c-Src. Proc Natl Acad Sci USA 105: 15914-15919. doi: 10.1073/pnas.0808171105
|
| [64] | Kumar K, Sachdanandam P, Arivazhagan R (1991) Studies on the changes in plasma lipids and lipoproteins proteins in patients with benign and malignant breast cancer. Biochem Int 23: 581-589. |
| [65] |
Damen J, Ramshorst JV, Hoeven RPV, et al. (1984) Alterations in plasma lipoprotein and heparin-releasable lipase activities in mice bearing the grsl ascites tumor. Biochim Biophys Acta 793: 287-296. doi: 10.1016/0005-2760(84)90331-X
|
| [66] |
Rengarajan T, Nandakumar N, Balasubramanian MP (2012) D-pinitol a low molecular cyclitol prevents 7, 12-dimethylbenz (a) anthracene induced experimental breast cancer through regulating anti-apoptotic protein Bcl-2, mitochondrial and carbohydrate key metabolizing enzymes. Biomed Prev Nutr 2: 25-30. doi: 10.1016/j.bionut.2011.11.001
|
| [67] |
Rawal LB, Tapp RJ, Williams ED, et al. (2012) Prevention of type 2 diabetes and its complications in developing countries: A review. Int J Behav Med 19: 121-133. doi: 10.1007/s12529-011-9162-9
|
| [68] |
Rathmann W, Giani G (2004) Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care 27: 1047-1053. doi: 10.2337/diacare.27.10.2568
|
| [69] |
Zhang BB, Moller DE (2000) New approaches in the treatment of type 2 diabetes. Curr Opin Chem Biol 4: 461-467. doi: 10.1016/S1367-5931(00)00103-4
|
| [70] |
Buse JB (2011) Type 2 diabetes mellitus in 2010: Individualizing treatment targets in diabetes care. Nat Rev Endocrinol 7: 67-68. doi: 10.1038/nrendo.2010.230
|
| [71] |
Sivakumar S, Palsamy P, Subramanian SP (2010) Impact of D-pinitol on the attenuation of proinflammatory cytokines, hyperglycemia-mediated oxidative stress and protection of kidney tissue ultrastructure in streptozotocin-induced diabetic rats. Chem Biol Interact 188: 237-245. doi: 10.1016/j.cbi.2010.07.014
|
| [72] |
Geethan PK, Prince PS (2008) Antihyperlipidemic effect of D-pinitol on streptozotocin-induced diabetic Wistar rats. J Biochem Mol Toxicol 22: 220-224. doi: 10.1002/jbt.20218
|
| [73] | WHO Expert Committee on Diabetes Mellitus: Second Report (1980) World Health Organ Tech Rep Ser 646: 1-80. |
| [74] |
Bates SH, Jones RB, Bailey CJ (2000) Insulin like effect of pinitol. Br J Pharmacol 130: 1944-1948. doi: 10.1038/sj.bjp.0703523
|
| [75] | Narayanan CR, Joshi DD, Mujumdar AM, et al. (1987) Pinitol, a new antidiabetic compound from the leaves of Bougainvillea pectabilis. Curr Sci 56: 139-141. |
| [76] | Rastegar S, Soltani S, Roohipoor A, et al. (2017) Study of plants with D-chiro-inositol and its derivatives on diabetes. Int J Pharmacogn 4: 43-53. |
| [77] |
Nascimento N, Cortez LU, Sousa LG, et al. (2014) Pinitol ameliorates impaired pressurenatriuresis in experimental diabetes. FASEB J 28: 1063-1065. doi: 10.1096/fasebj.28.1_supplement.1063.5
|
| [78] |
Sivakumar S, Palsamy P, Subramanian SP (2010) Attenuation of oxidative stress and alteration of hepatic tissue ultrastructure by D-pinitol in streptozotocin-induced diabetic rats. Free Radic Res 44: 668-678. doi: 10.3109/10715761003733901
|
| [79] |
Sivakumar S, Subramanian SP (2009) Pancreatic tissue protective nature of D-pinitol studied in streptozotocin-mediated oxidative stress in experimental diabetic rats. Eur J Pharmacol 622: 65-70. doi: 10.1016/j.ejphar.2009.09.021
|
| [80] |
Davis A, Christiansen M, Horowitz JF, et al. (2000) Effect of pinitol treatment on insulin action in subjects with insulin resistance. Diabetes Care 23: 1000-1005. doi: 10.2337/diacare.23.7.1000
|
| [81] |
Ortmeyer HK, Huang LC, Zhang L, et al. (1993) Chiroinositol deficiency and insulin resistance. Ⅱ. Acute effects of D-chiroinositol administration in streptozotocin diabetic rats, normal rats given a glucose load, and spontaneously insulin-resistant rhesus monkeys. Endocrinology 132: 646-651. doi: 10.1210/endo.132.2.8425484
|
| [82] |
Fonteles MC, Huang LC, Larner J (1996) Infusion of pH 2.0 D-chiroinositol glycan insulin putative mediator normalizes plasma glucose in streptozotocin diabetic rats at a dose equivalent to insulin without inducing hypoglycaemia. Diabetologia 39: 731-734. doi: 10.1007/BF00418546
|
| [83] |
Fonteles MC, Almeida MQ, Larner J (2000) Antihyperglycemic effects of 3-O-methyl-D-chiro-inositol and D-chiro-inositol associated with manganese in sterptozotocin diabetic rats. Horm Metab Res 32: 129-132. doi: 10.1055/s-2007-978606
|
| [84] |
Ortmeyer HK, Larner J, Hansen BC (1995) Effects of D-chiroinositol added to a meal on plasma glucose and insulin in hyperinsulinemic rhesus monkeys. Obes Res 3: 605S-608S. doi: 10.1002/j.1550-8528.1995.tb00232.x
|
| [85] |
Kang MJ, Kim JI, Yoon SY, et al. (2006) Pinitol from soybeans reduces postprandials blood glucose in patients with type 2 diabetes mellitus. J Med Food 9: 182-186. doi: 10.1089/jmf.2006.9.182
|
| [86] |
Dang NT, Mukai R, Yoshida K, et al. (2010) D-pinitol and myo-inositol stimulate translocation of glucose transporter 4 in skeletal muscle of C57BL/6 mice. Biosci Biotechnol Biochem 74: 1062-1067. doi: 10.1271/bbb.90963
|
| [87] |
Yu J, Choi S, Park ES, et al. (2012) D-chiro-inositol negatively regulates the formation of multinucleated osteoclasts by down-regulating NFATc1. J Clin Immunol 32: 1360-1371. doi: 10.1007/s10875-012-9722-z
|
| [88] |
Kim JI, Kim JC, Kang MJ, et al. (2005) Effects of pinitol isolated from soybeans on glycemic control and cardiovascular risk factors in patients with type 2 diabetes mellitus: A randomized controlled study. Eur J Clin Nutr 59: 456-458. doi: 10.1038/sj.ejcn.1602081
|
| [89] |
Kim MJ, Yoo KH, Kim JH, et al. (2007) Effect of pinitol on glucose metabolism and adipocytokines in uncontrolled type 2 diabetes. Diabetes Res Clin Pract 77: S247-S251. doi: 10.1016/j.diabres.2007.01.066
|
| [90] |
Kim HJ, Park KS, Lee SK, et al. (2012) Effects of pinitol on glycemic control, insulin resistance and adipocytokine levels in patients with type 2 diabetes mellitus. Ann Nutr Metab 60: 1-5. doi: 10.1159/000334834
|
| [91] |
Yap A, Nishiumi S, Yoshida KI, et al. (2007) Rat L6 myotubes as an in vitro model system to study GLUT4-dependent glucose uptake stimulated by inositol derivatives. Cytotechnology 55: 103-108. doi: 10.1007/s10616-007-9107-y
|
| [92] |
Larner J (2002) D-chiro-inositol its functional role in insulin action and its deficit in insulin resistance. Int J Exp Diabetes Res 3: 47-60. doi: 10.1080/15604280212528
|
| [93] |
Yamashita Y, Yamaoka M, Hasunuma T, et al. (2013) Detection of orally administered inositol stereoisomers in mouse blood plasma and their effects on translocation of glucose trasporter 4 in skeletal muscle cells. J Agric Food Chem 61: 4850-4854. doi: 10.1021/jf305322t
|
| [94] |
Hernández-Mijares A, Bañuls C, Peris JE, et al. (2013) A single acute dose of pinitol from a naturally-occurring food ingredient decreases hyperglycaemia and circulating insulin levels in healthy subjects. Food Chem 141: 1267-1272. doi: 10.1016/j.foodchem.2013.04.042
|
| [95] | Martin-Lomas M, Rademacher TW, Caro HN, et al. (2001) Alkylated inositolglycans and their use. Worldwide patent WO 0185747(A1). |
| [96] | Ostlund RE, Sherman WR (1998) Pinitol and derivatives thereof for the treatment of metabolic disorders. United States patent US 5827896(A). |
| [97] | Larner J, Price J, Picariello T, et al. (1997) Method of treating defective glucose metabolism using synthetic insulin substances. United States patent US 5652221(A). |
| [98] | Koon MH (2013) Combination of pinitol and natural product for treating diabetes mellitus. Korean patent KR 20130017864(A). |
| [99] | Koon MH (2013) Combination of pinitol and drug for treating diabetes mellitus. Korean patent KR 20130017859(A). |
| [100] | Jun JG, Jun YJ, Kim JJ, et al. (2004) Use of chiro-inositol or pinitol for prevention of oxidative damage and prophylaxis composition for diabetic complications containing the chiro-inositol or pinitol. Korean patent KR 20040051455(A). |
| [101] |
Holman GD, Kasuga M (1997) From receptor to transporter: Insulin signalling to glucose transport. Diabetologia 40: 991-1003. doi: 10.1007/s001250050780
|
| [102] |
White MF (1997) The insulin signaling system and IRS proteins. Diabetologia 40: S2-S17. doi: 10.1007/s001250051387
|
| [103] |
Huang LC, Fonteles MC, Houston DB, et al. (1993) Chiroinositol deficiency and insulin resistance. Ⅲ. Acute glycogenic and hypoglycemic effects of two inositol phospsoglycan insulin mediators in normal and streptozotocin diabetic rats. Endocrinology 132: 652-657. doi: 10.1210/endo.132.2.8425485
|
| [104] | PI3 Kinase/Akt Signaling Pathway, In: Cell Signaling Technology. Available from: https://www.cellsignal.com/contents/science-pathway-research-pi3k-akt-signaling-resources/pi3k-akt-signaling-pathway/pathways-akt-signaling. |
| [105] |
Rahal A, Kumar A, Singh V, et al. (2014) Oxidative stress, pro-oxidants, and antioxidants: The interplay. Biomed Res Int 2014: 761264. doi: 10.1155/2014/761264
|
| [106] | Shin YC, Jeon YJ, Kim JJ (2007) Use of pinitol or chiroinositol for protecting the liver. United States patent US 2007098826 (A1). |
| [107] |
Magielse J, Arcoraci T, Breynaert A, et al. (2013) Antihepatotoxic activity of a quantified desmodium adscendens decoction and D-pnitol against chemically-induced liver in rats. J Ethnopharmacol 146: 250-256. doi: 10.1016/j.jep.2012.12.039
|
| [108] |
Keppler D, Lesch R, Reutter W, et al. (1968) Experimental hepatitis induced by D-galactosamine. Exp Mol Pathol 9: 279-290. doi: 10.1016/0014-4800(68)90042-7
|
| [109] |
Beck G, Habicht GS (1996) Immunity and the invertebrates. Sci Am 275: 60-66. doi: 10.1038/scientificamerican1196-60
|
| [110] | Alexander P (1975) Tumour immunology in perspective, In: Schcultz J, Leiff RC. Eds., Critical factors in cancer immunology, New York: Academic Press, 213-222. |
| [111] |
Katz A (1983) Immunobiologic staging of patients with carcinoma of the nad and neck. Laryngoscope 93: 445-463. doi: 10.1002/lary.1983.93.4.445
|
| [112] |
Gleich GJ, kita H (1997) Bronchial asthma: Lessons from murine models. Proc Natl Acad Sci USA 94: 2101-2102. doi: 10.1073/pnas.94.6.2101
|
| [113] |
Chauhan PS, Gupta KK, Bani S (2011) The immunosuppressive effects of Agyrolobium roseum and pinitol in experimental animals. Int Immunopharmacol 11: 286-291. doi: 10.1016/j.intimp.2010.11.028
|
| [114] |
Kim JC, Shin JY, Shin DH, et al. (2005) Synergistic anti-inflammatory effects of pinitol and glucosamine in rats. Phytother Res 19: 1048-1051. doi: 10.1002/ptr.1788
|
| [115] |
Lee JS, Lee CM, Jeong YI, et al. (2007) D-pinitol regulates Th1/Th2 balance via suppressing Th2 immune response in ovalbumin-induced asthma. FEBS Lett 581: 57-64. doi: 10.1016/j.febslet.2006.11.077
|
| [116] | Lee JS, Jung ID, Jeong YI, et al. (2007) D-pinitol inhibits Th1 polarization via the suppression of dendritic cells. Int Immunopharmacol 7: 79-804. |
| [117] | Heo JC, Lee SH, Hwang YH, (2011) Extract of immuno-suppressive activities of pinitol isolated from soybean. Korean patent KR 20110116627(A). |
| [118] | Yun YC, Choi CM, Jeon YJ, (2007) Composition for preventing and treating inflammatory disease comprising glucosamine and Pinitol. Korean patent KR 20070002401(A). |
| [119] |
Goltzman D (2002) Discoveries, drugs and skeletal disorders. Nat Rev Drug Discov 1: 784-796. doi: 10.1038/nrd916
|
| [120] |
Rodan GA, Martin TJ (2000) Therapeutic approaches to bone diseases. Science 289: 1508-1514. doi: 10.1126/science.289.5484.1508
|
| [121] | Jaerang R, Hyeon-Koon M (2010) Composition for prevention or treatment of bone metabolism disorder comprising d-pinitol as an active ingredient. Chinese patent CN 101808628(A). |
| [122] |
Kirkwood TB, Austad SN (2000) Why do we age? Nature 408: 233-238. doi: 10.1038/35041682
|
| [123] |
Hada B, Yoo MR, Seong KM, et al. (2013) D-chiro-inositol and pinitol extend the life span of Drosophila Melanogaster. J Gerontol 68: 226-234. doi: 10.1093/gerona/gls156
|
| [124] | Fontana L, Partridge L, Longo V (2010) Science 328: 321-326. |
| [125] |
Bartke A, Chandrashekar V, Dominici F, et al. (2003) Insulin-like growth fact 1 (IGF-1) and aging: Controversies and new insights. Biogerontology 4: 1-8. doi: 10.1023/A:1022448532248
|
| [126] | Van-Heemst D (2010) Insulin, IGF-1 and logevity. Aging Dis 1: 147-157. |
| [127] |
Chung HY, Kim HJ, Kim JW, et al. (2001) The inflammation hypothesis of aging: Molecular modulation by calorie restriction. Ann N Y Acad Sci 928: 327-335. doi: 10.1111/j.1749-6632.2001.tb05662.x
|
| [128] | Ishida M, Suzuki T, Tsuji A (2015) Biological clock adjusting agent. Japanese patent JP 2015140305(A). |
| [129] | Choi SK, Park KD, Kim DA, et al. (2013) Preparation method for Ceratonia siliqua fruit extract and cosmetic composition for anti-aging comprising the same. Korean patent KR 101339915(B1). |
| [130] | Choi SK, Park KD, Kim DA, et al. (2015) Cosmetic composition for anti-aging comprising Ceratonia siliqua fruit extract. Korean patent KR 20150060004(A). |
| [131] |
Barcelos RP, Stefanello ST, Mauriz JL, et al. (2016) Creatine and the liver: Metabolism and possible interactions. Mini Rev Med Chem 16: 12-18. doi: 10.2174/1389557515666150722102613
|
| [132] |
Cooper R, Naclerio F, Allgrove J, et al. (2012) Creatine supplementation with specific view to exercise/sports performance: An update. J Int Soc Sports Nutr 9: 1-11. doi: 10.1186/1550-2783-9-33
|
| [133] | Greenwood M, Kreider RB, RasMussen C, et al. (2001) D-pinitol augments whole body creatine retention in man. J Exerc Physiol Online 4: 41-47. |
| [134] | Cooke MB, Cribb PJ (2015) Effective nutritional supplement combinations, In: Greenwood M, Cooke MC, Ziegenfuss T, Kalman DS, Jose-Antonio Eds., Nutritional Supplements in Sports and Exercise, 2nd ed., Switzerland: Springer. |
| [135] | Chantler S, Smit K (2015) Pinitol, In: Castell LM, Stear SJ, Burke LM Eds., Nutritional Supplements in Sport, Exercise and Health: An A-Z Guide, London and New York: Routledge-Taylor & Francis Group. |
| [136] |
Kerksick CM, Wilborn CD, Campbell WI, et al. (2009) The effects of creatine monohydrate supplementation with and without D-pinitol on resistance training adaptations. J Strength Cond Res 23: 2673-2682. doi: 10.1519/JSC.0b013e3181b3e0de
|
| [137] | Dykstra JC (2001) A combination of pinitol and creatine to enhance uptake and retention of creatine. Worldwide patent WO 0180853(A1). |
| [138] | Heuer MA, Clement K, Chaudhuri S (2008) Composition and method for enhancing or promoting the activity of insulin, enhancing skeletal muscle growth, reducing skeletal muscle loss, and increasing the energy supply to skeletal muscle. Worldwide patent WO 2008025116(A1). |
| [139] | Weeks C (2003) Stimulating transport of glucose into animal administration of pinitol. United States patent US 6518318. |
| [140] | Ferrante RM, Cunnigham CK (2012) Performance enhancing composition and method of delivering nutrients. United States patent US 2012100247(A1). |
| [141] | Ferrante RM, Cunnigham CK (2012) Performance enhancing compositions and method of delivering nutrients. United States patent US 2012100120(A1). |
| [142] | Ferrante RM, Cunnigham CK (2015) Performance enhancing composition and method of delivering nutrients. United States patent US 2015196579(A1). |
| [143] | Heuer MA, Clement K, Chaudhuri S (2008) Composition and method for enhancing or promoting the activity of insulin, enhancing skeletal muscle growth, reducing skeletal muscle loss, and increasing the energy supply to skeletal muscle. United States patent US 2008058254(A1). |
| [144] | Dykstra JC, Prairie E (2003) Combination of pinitol and creatine to enhance uptake and retention of creatine. United States patent US 2003212134(A1). |
| [145] | Heuer MA, Clement K, Chaudhuri S (2008) Composition and method for enhancing or promoting the activity of insulin, enhancing skeletal muscle growth, reducing skeletal muscle loss, and increasing the energy supply to skeletal muscle. Canadian patent CA 2558110(A1). |
| [146] |
Folch J, Petrov D, Ettcheto M, et al. (2016) Current research therapeutic strategies for Alzheimer's disease treatment. Neural Plast 2016: 1-15. doi: 10.1155/2016/8501693
|
| [147] |
Pitt J, Thorner M, Brautigan D, et al. (2013) Protection against the synaptic targeting and toxicity of Alzheimer's-associated Aβ oligomers by insulin mimetic chiro-inositols. FASEB J 27: 199-207. doi: 10.1096/fj.12-211896
|
| [148] |
Wischik CM, Harrington CR, Storey JMD (2014) Tau-aggregation inhibitor therapy for Alzheimer's disease. Biochem Pharmacol 88: 529-539. doi: 10.1016/j.bcp.2013.12.008
|
| [149] |
Shea TB, Remington R (2015) Nutritional supplementation for Alzheimer's disease? Curr Opin Psychiatry 28: 141-147. doi: 10.1097/YCO.0000000000000138
|
| [150] |
Amirrad F, Bousoik E, Shamloo K, et al. (2017) Alzheimer's disease: Dawm of a new era? J Pharm Pharm Sci 20: 184-225. doi: 10.18433/J3VS8P
|
| [151] |
Hung SY, Fu WM (2017) Drug candidates in clinical trials for Alzheimer's disease. J Biomed Sci 24: 1-12. doi: 10.1186/s12929-016-0310-z
|
| [152] | Acton QA (2013) Therapies and treatments, In: Neurodegenerative diseases: New insights for the healthcare professional, Georgia: ScholarlyEditions, 203-204. |
| [153] | Yates P, Woodward M (2017) Drug treatments in development for Alzheimer's disease, In: Ames D, O'Brien JT, Burns A. Editors, Dementia, 5 Eds., New York: CRC Press, 559. |
| [154] | Pasinetti GM (2006) Compositions and methods for treating Alzheimer's disease and related disorders and promoting a healthy nervous system. United States patent US 2006/0111450A1. |
| [155] | Pasinetti GM (2013) Compositions and methods for treating Alzheimer's disease and related disorders and promoting a healthy nervous system. United States patent US 2013/0123370A1. |
| [156] | McLaurin J (2010) Methods of preventing, treating and diagnosing disorders of protein aggregation. European patent EU 2153829A1. |
| [157] | Available from: a) https://clinicaltrials.gov/ct2/show/NCT00470418. b) Available from: https://clinicaltrials.gov/ct2/show/NCT01928420. |
| [158] |
Falshaw A, Hart JB, Tyler PC (2000) New synthesis of 1 d-and 1 L-1, 2-anhydro-myo-inositol and assessment of their glycosidase inhibitory activities. Carbohydr Res 329: 301-308. doi: 10.1016/S0008-6215(00)00192-0
|
| [159] |
Zhan T, Lou H (2007) Synthesis of azole nucleoside analogues of D-pinitol as potential antitumor agents. Carbohydr Res 342: 865-869. doi: 10.1016/j.carres.2007.01.004
|
| [160] |
Li M, Wu A, Zhou P (2006) A concise synthesis of (+)-pancratistatin using pinitol as a chiral building blog. Tetrahedron Lett 47: 3707-3710. doi: 10.1016/j.tetlet.2006.03.138
|
| [161] |
Tegge W, Ballou CE (1989) Chiral synthesis of D-and L-myo-inositol 1, 4, 5-triphosphate. Proc Natl Acad Sci USA 86: 94-98. doi: 10.1073/pnas.86.1.94
|
| [162] |
Ballou CE, Fischer HOL (1953) Derivatives of D-mannohexodialdose (6-aldo-D-Mannose). J Am Chem Soc 75: 3673-3675. doi: 10.1021/ja01111a020
|
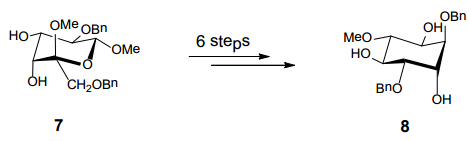
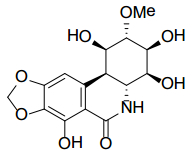
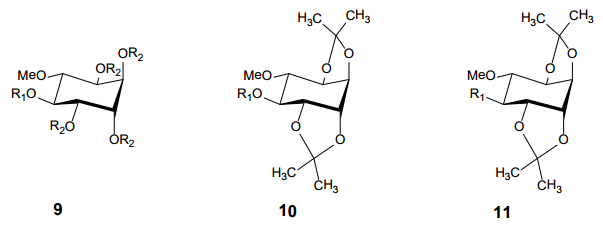
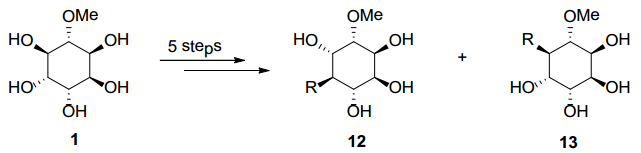

Figures(9) / Tables(2)

JoséIgnacio López-Sánchez, Diego A. Moreno, Cristina García-Viguer. D-pinitol, a highly valuable product from carob pods: Health-promoting effects and metabolic pathways of this natural super-food ingredient and its derivatives[J]. AIMS Agriculture and Food, 2018, 3(1): 41-63. doi: 10.3934/agrfood.2018.1.41







 DownLoad:
DownLoad: