Citation: Jin Liang, Hong-Ming Yin, Xinfu Chen, Yuan Wu. On a Corporate Bond Pricing Model with Credit Rating Migration Risksand Stochastic Interest Rate[J]. Quantitative Finance and Economics, 2017, 1(3): 300-319. doi: 10.3934/QFE.2017.3.300

| [1] |
Black F, Cox JC (1976) Some Effects of Bond Indenture Provisions. J Financ 31: 351-367. doi: 10.1111/j.1540-6261.1976.tb01891.x

|
| [2] |
Briys E, Varenne DF (1997) Valuing Risky Fixed Rate Debt: An Extension. J Financ Quantit Anal 32: 239-249. doi: 10.2307/2331175
|
| [3] |
Duffe D, Singleton KJ (1999) Modeling Term Structures of Defaultable Bonds. Rev Financ Stud 12: 687-720. doi: 10.1093/rfs/12.4.687
|
| [4] | Das S, Tufano P (1996) Pricing credit-sensitive debt when interest rates, credit ratings, and credit spreads are stochastic. J Financ Eng 5: 161-198. |
| [5] | Dixit AK, Pindyck S (1994) Investment under Uncertainty, Princeton Univ Press. |
| [6] |
Evans LC (1978) A free boundary problem: the flow of two immiscible fluids in a one-dimensional porous medium. II. Indiana Univ Math J 27: 93-101. doi: 10.1512/iumj.1978.27.27009
|
| [7] | Friedman A (1982) Variational Principles and Free Boundary Problems. John Wiley Sons, New York. |
| [8] | Garrori MG; Menaldi JL (1992) Green Functions for Second Order Parabolic Integro-differential Problems. Longman Sci Tech, New York. |
| [9] | Hall J (1989) Options, Futures, and Other Derivatives, Prentice-Hall. Inc, New Jersey. |
| [10] | Hu B (2011) Blow-up Theories for Semilinear Parabolic Equations, Heidelberg ; New York : Springer. |
| [11] | Jiang L (2005) Mathematical Modeling and Methods for Option Pricing. World Sci. |
| [12] |
Hu B, Liang J, Wu Y (2015) A Free Boundary Problem for Corporate Bond with Credit Rating Migration. J Math Anal Appl 428: 896-909. doi: 10.1016/j.jmaa.2015.03.040
|
| [13] | Hurd T, Kuznetsov A (2007) Affine Markov chain models of multifirm credit migration. J Credit Risk 3: 3-29. |
| [14] |
Jarrow R, Turnbull S (1995) Pricing Derivatives on Financial Securities Subject to Credit Risk. J Financ 50: 53-86. doi: 10.1111/j.1540-6261.1995.tb05167.x
|
| [15] |
Jarrow RA, Lando D, TurnbullS M (1997) A Markov model for the term structure of credit risk spreads. Rev Financ stud 10: 481-523. doi: 10.1093/rfs/10.2.481
|
| [16] | Lando D (1998) On Cox Processes and Credit-risky Securities. Rev Deriv Res, 1998, 2:99-120. |
| [17] | Lando D (2000) Some elements of rating based credit risk modeling. Adv Fixed-Income Valuat Tools 193-215. |
| [18] | Leland H, Toft KB (1996) Optimal capital structure,endogenous bankruptcy,and the term strcuture of credit spreads. J Financ 3: 987-1019. |
| [19] |
Longstaff F, Schwartz E (1995) A Simple Approach to Valuing Risky Fixed and Floating Rate Debt. J Financ 50: 789-819. doi: 10.1111/j.1540-6261.1995.tb04037.x
|
| [20] | Ladyzenskaja OA, Solonnikov VA, Uralceva NN (1968) Linear and Quasilinear Equations of Parabolicn Type. AMS Transl Math Monogr 23. |
| [21] | Liang J, Wu Y, Hu B (2016) Asymptotic Traveling Wave Solution for a Credit Rating Migration Problem. J Differ Equ 261: 1017-1045. |
| [22] | Liang J, Zeng ZK (2015) Pricing on Corporate Bonds with Credit Rating Migration under Structure Framework, Appl Math A J Chin Univ. |
| [23] | Liang J, Zhao YJ (2014) Utility Indifference Valuation of Corporate Bond with Credit Rating Migration by Structure Approach, preprint. |
| [24] | Merton RC (1974) On the Pricing of Corporate Debt: The Risk Structure of Interest Rates. J Financ 29: 449-470. |
| [25] |
Thomas L, Allen D, Morkel-Kingsbury N (2002) A hidden Markov chain model for the term structure of bond credit risk spreads. Int Rev Financ Anal 11: 311-329. doi: 10.1016/S1057-5219(02)00078-9
|
| [26] |
Tsiveriotis K, Fernandes C (1998) Valuing convertible bonds with credit risk. J Fixed Income 8: 95-102. doi: 10.3905/jfi.1998.408243
|
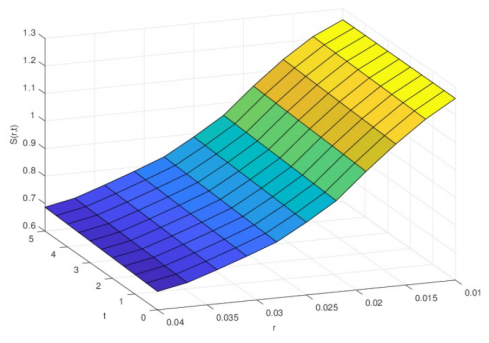
Figures(1)

Jin Liang, Hong-Ming Yin, Xinfu Chen, Yuan Wu. On a Corporate Bond Pricing Model with Credit Rating Migration Risksand Stochastic Interest Rate[J]. Quantitative Finance and Economics, 2017, 1(3): 300-319. doi: 10.3934/QFE.2017.3.300







 DownLoad:
DownLoad: