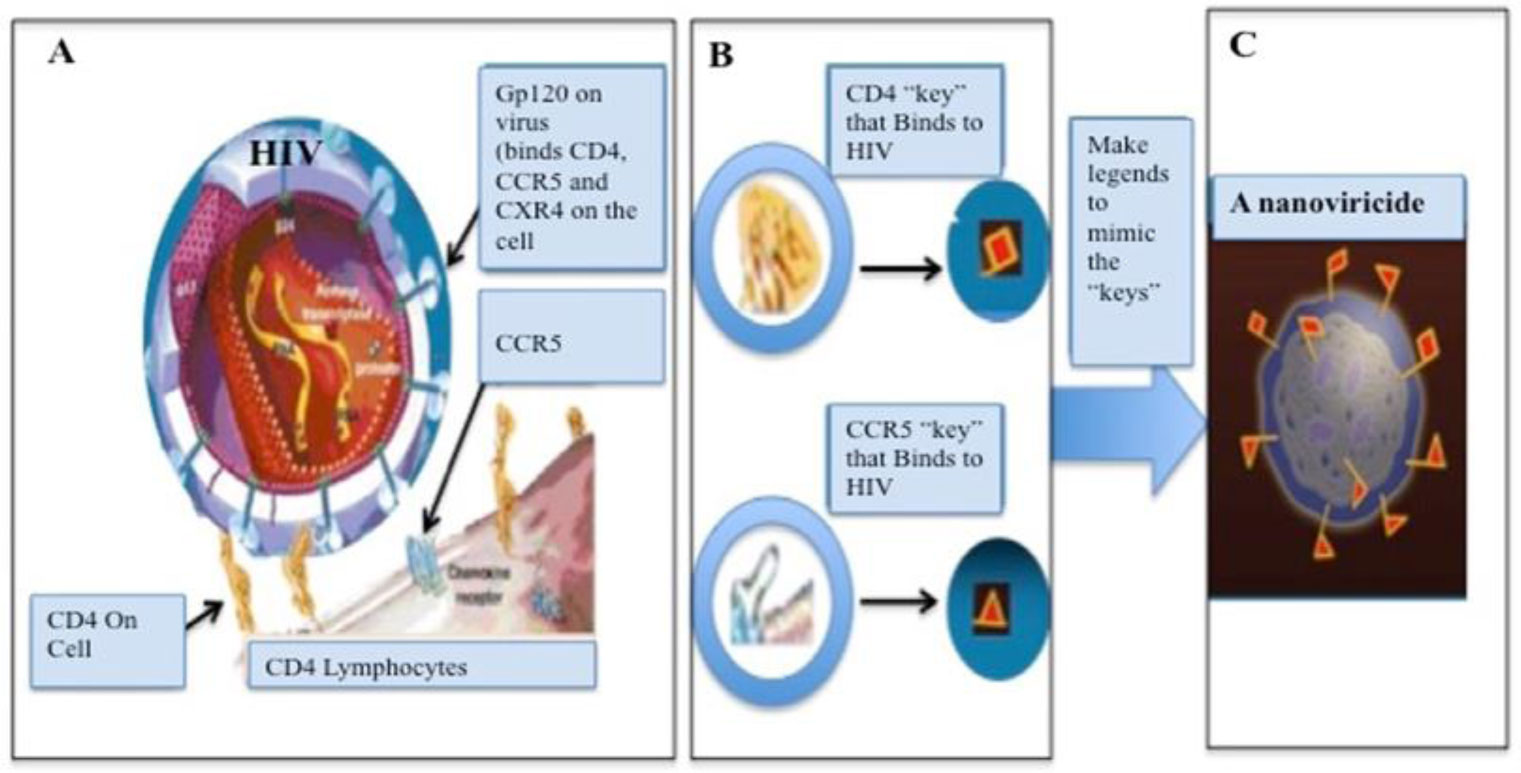
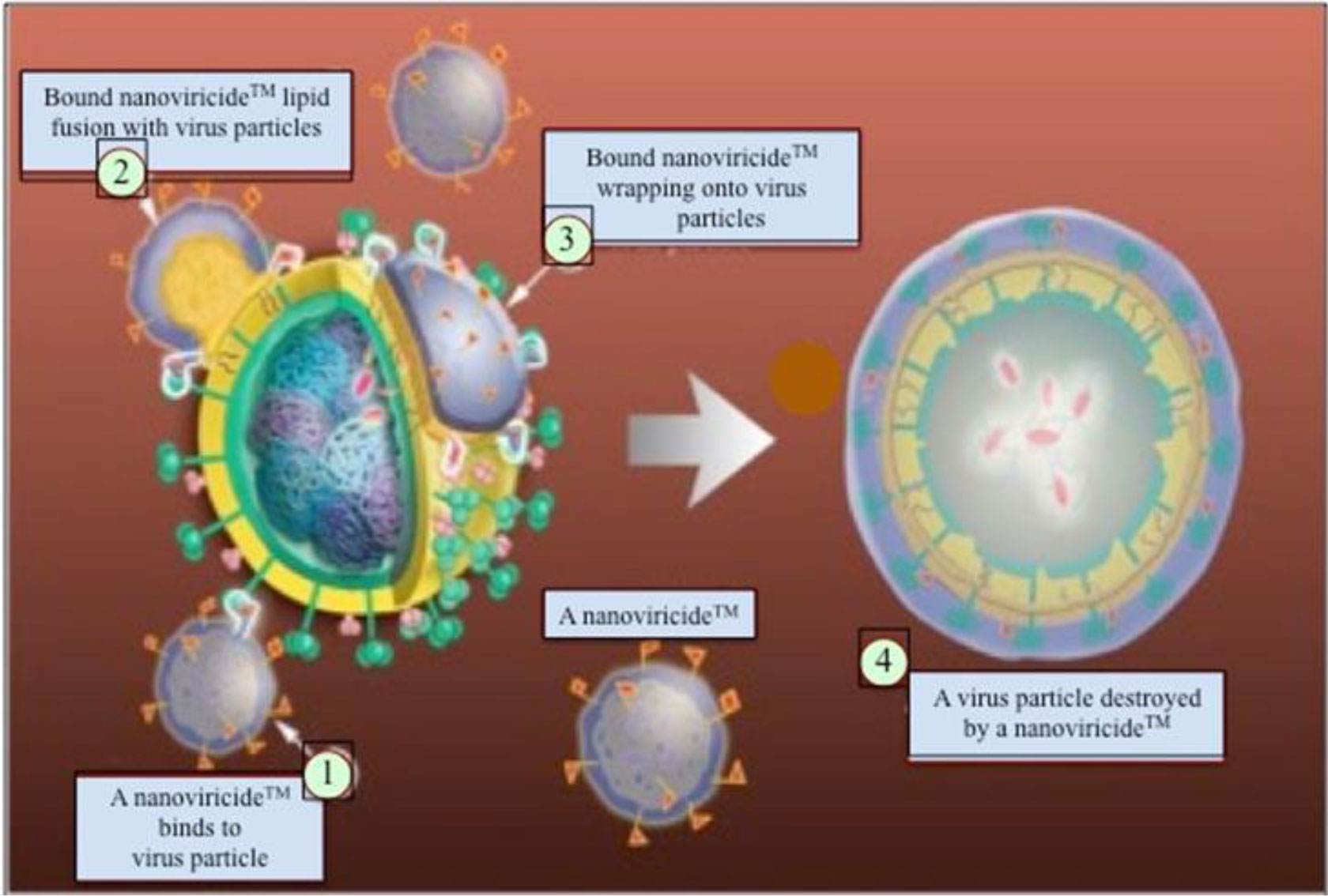
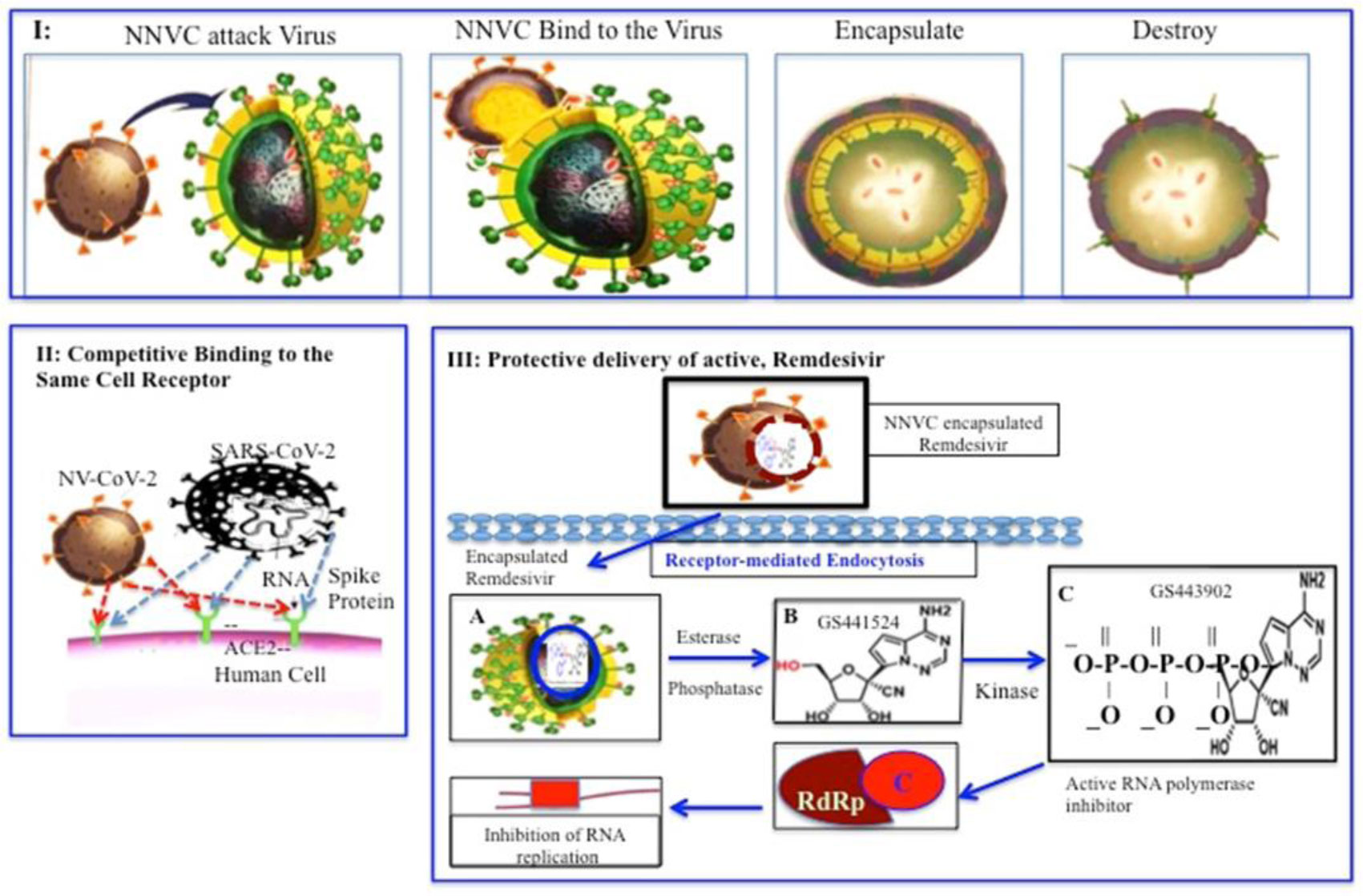
NV-CoV-2 is a nanoviricide that is covalently bonded with polyethylene glycol (PEG) and alkyl pendants. This molecular design is used to attack many strains of coronaviruses in a broad-spectrum manner. The ligand works by competitive inhibition and binds to the same site on the S-protein of SARS-CoV that attaches to the cognate cellular receptor, ACE2. This prevents SARS-CoV from binding and infecting the cell. NV-CoV-2 is designed to bind to the free virion particles at multiple points encapsulate the virus and disable its ability to infect the cells. The multi-point binding interaction, like a nano-velcro-tape, may lead to lipid-lipid fusion of the alkyl chains in the nanoviricide micelle with the lipid envelope of the virus. The virus becomes dismantled to a capsid form before the host immune system becomes involved. This putative mechanism is orthogonal to many other anti-coronavirus agents in development. Thus, it maybe possible to produce a stronger antiviral effect when combining NV-CoV-2 therapy with other anti-coronavirus therapies such as Remdesivir (RDV). NV-CoV-2 can encapsulate other antiviral compounds as well. In this study, RDV was encapsulated and protected from serum-mediated degradation in vivo. As a result, RDV was available for a longer period of time to interact with RNA polymerase and inhibit.
Citation: Ashok Chakraborty, Anil Diwan, Vinod Arora, Yogesh Thakur, Vijetha Chiniga, Jay Tatake, Rajesh Pandey, Preetam Holkar, Neelam Holkar, Bethany Pond. Mechanism of Antiviral Activities of Nanoviricide's Platform Technology based Biopolymer (NV-CoV-2)[J]. AIMS Public Health, 2022, 9(2): 415-422. doi: 10.3934/publichealth.2022028
NV-CoV-2 is a nanoviricide that is covalently bonded with polyethylene glycol (PEG) and alkyl pendants. This molecular design is used to attack many strains of coronaviruses in a broad-spectrum manner. The ligand works by competitive inhibition and binds to the same site on the S-protein of SARS-CoV that attaches to the cognate cellular receptor, ACE2. This prevents SARS-CoV from binding and infecting the cell. NV-CoV-2 is designed to bind to the free virion particles at multiple points encapsulate the virus and disable its ability to infect the cells. The multi-point binding interaction, like a nano-velcro-tape, may lead to lipid-lipid fusion of the alkyl chains in the nanoviricide micelle with the lipid envelope of the virus. The virus becomes dismantled to a capsid form before the host immune system becomes involved. This putative mechanism is orthogonal to many other anti-coronavirus agents in development. Thus, it maybe possible to produce a stronger antiviral effect when combining NV-CoV-2 therapy with other anti-coronavirus therapies such as Remdesivir (RDV). NV-CoV-2 can encapsulate other antiviral compounds as well. In this study, RDV was encapsulated and protected from serum-mediated degradation in vivo. As a result, RDV was available for a longer period of time to interact with RNA polymerase and inhibit.

| [1] | Diwan A, Chakraborty A, Tatake J, et al. (2021) Broad spectrum antiviral therapy can supersede vaccination strategy for combating COVID-19 pandemic. Med J Clin Trials Case Stud 5: 298-304. https://doi.org/10.23880/mjccs-16000298 |
| [2] | Diwan A, Chakraborty A, Chiniga V, et al. (2021) Dual Effects of Nanoviricides Platform Technology Based NV-CoV-2 Biomimetic Polymer Against COVID-19. BioRxiv . https://doi.org/10.1101/2021.11.24.469813 |
| [3] | Chakraborty A, Diwan A, Arora V, et al. (2022) Nanoviricides Platform Technology Based NV-387 Polymer Protects Remdesivir from Plasma-Mediated Catabolism In vitro: Importance of its Increased Lifetime for In vivo Action. Recent Adv Clin Trials 1: 1-8. |
| [4] | Chakraborty A, Diwan A, Arora V, et al. (2021) Nanoviricide's platform technology based NV-CoV-2 polymer increases the half-life of Remdesivir in vivo. BioRxiv . https://doi.org/10.1101/2021.11.17.468980 |
| [5] | Chakraborty A, Diwan A, Arora V, et al. (2022) Encapsulation of Remdesivir in Nanoviricide's Platform Technology Based NV-CoV-2 Polymer Protects the Drug and Improves Its Pharmacokinetics. EC Pharmacol Toxicol 10.2: 108-118. |
| [6] |
Li W, Sui J, Huang IC, et al. (2007) The S proteins of human coronavirus NL63 and severe acute respiratory syndrome coronavirus bind overlapping regions of ACE2. Virology 367: 367-374. https://doi.org/10.1016/j.virol.2007.04.035 
|
| [7] |
Chakraborty A, Diwan A (2020) NL-63: A better surrogate virus for studying SARS-CoV-2. Integr Mol Med 7: 1-9. https://doi.org/10.15761/IMM.1000408
|
| [8] |
Hofmann H, Pyrc K, Van Der Hoek L, et al. (2005) Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci USA 102: 7988-7993. https://doi.org/10.1073/pnas.0409465102
|
| [9] | Liu DX, Liang JQ, Fung TS (2021) Human Coronavirus-229E, -OC43, -NL63, and -HKU1 (Coronaviridae). Encycl Virol : 428-440. https://doi.org/10.1016/B978-0-12-809633-8.21501-X |
| [10] |
Belouzard S, Millet JK, Licitra BN, et al. (2012) Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 4: 1011-1033. https://doi.org/10.3390/v4061011
|
| [11] |
Chakraborty A, Diwan A (2020) Pharmacodynamics of Remdesivir: How to improve for COVID-19 Treatment. J Biomed Res Environ Sci 1: 431-438. https://doi.org/10.37871/jbres1175
|
| [12] | Barton RW, Tatake JG, Diwan AR (2016) Nanoviricides: Targeted Anti-Viral Nanomaterials. Handbook of Clinical Nanomedicine, Nanoparticles, Imaging, Therapy, and Clinical Applications . Jenny Stanford Publishing 1039-1046. |
| [13] | Barton RW, Tatake JG, Diwan AR (2011) Nanoviricides-A Novel Approach to Antiviral Therapeutics. Bionanotechnology II. Ed. 160-173. https://doi.org/10.1201/b11374-11 |

Figures(4)

Ashok Chakraborty, Anil Diwan, Vinod Arora, Yogesh Thakur, Vijetha Chiniga, Jay Tatake, Rajesh Pandey, Preetam Holkar, Neelam Holkar, Bethany Pond. Mechanism of Antiviral Activities of Nanoviricide's Platform Technology based Biopolymer (NV-CoV-2)[J]. AIMS Public Health, 2022, 9(2): 415-422. doi: 10.3934/publichealth.2022028







 DownLoad:
DownLoad: