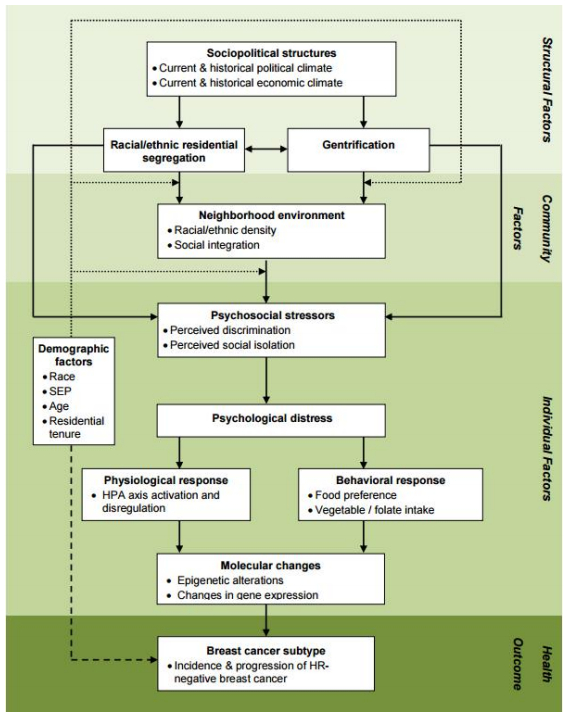
Hormone receptor negative (HR-) breast cancer subtypes are etiologically distinct from the more common, less aggressive, and more treatable form of estrogen receptor positive (ER+) breast cancer. Numerous population-based studies have found that, in the United States, Black women are 2 to 3 times more likely to develop HR- breast cancer than White women. Much of the existing research on racial disparities in breast cancer subtype has focused on identifying predisposing genetic factors associated with African ancestry. This approach fails to acknowledge that racial stratification shapes a wide range of environmental and social exposures over the life course. Human stress genomics considers the role of individual stress perceptions on gene expression. Yet, the role of structurally rooted biopsychosocial processes that may be activated by the social patterning of stressors in an historically unequal society, whether perceived by individual black women or not, could also impact cellular physiology and gene expression patterns relevant to HR- breast cancer etiology. Using the weathering hypothesis as our conceptual framework, we develop a structural perspective for examining racial disparities in breast cancer subtypes, integrating important findings from the stress biology, breast cancer epidemiology, and health disparities literatures. After integrating key findings from these largely independent literatures, we develop a theoretically and empirically guided framework for assessing potential multilevel factors relevant to the development of HR- breast cancer disproportionately among Black women in the US. We hypothesize that a dynamic interplay among socially patterned psychosocial stressors, physiological & behavioral responses, and genomic pathways contribute to the increased risk of HR- breast cancer among Black women. This work provides a basis for exploring potential alternative pathways linking the lived experience of race to the risk of HR- breast cancer, and suggests new avenues for research and public health action.
Citation: Erin Linnenbringer, Sarah Gehlert, Arline T. Geronimus. Black-White Disparities in Breast Cancer Subtype: The Intersection of Socially Patterned Stress and Genetic Expression[J]. AIMS Public Health, 2017, 4(5): 526-556. doi: 10.3934/publichealth.2017.5.526
Hormone receptor negative (HR-) breast cancer subtypes are etiologically distinct from the more common, less aggressive, and more treatable form of estrogen receptor positive (ER+) breast cancer. Numerous population-based studies have found that, in the United States, Black women are 2 to 3 times more likely to develop HR- breast cancer than White women. Much of the existing research on racial disparities in breast cancer subtype has focused on identifying predisposing genetic factors associated with African ancestry. This approach fails to acknowledge that racial stratification shapes a wide range of environmental and social exposures over the life course. Human stress genomics considers the role of individual stress perceptions on gene expression. Yet, the role of structurally rooted biopsychosocial processes that may be activated by the social patterning of stressors in an historically unequal society, whether perceived by individual black women or not, could also impact cellular physiology and gene expression patterns relevant to HR- breast cancer etiology. Using the weathering hypothesis as our conceptual framework, we develop a structural perspective for examining racial disparities in breast cancer subtypes, integrating important findings from the stress biology, breast cancer epidemiology, and health disparities literatures. After integrating key findings from these largely independent literatures, we develop a theoretically and empirically guided framework for assessing potential multilevel factors relevant to the development of HR- breast cancer disproportionately among Black women in the US. We hypothesize that a dynamic interplay among socially patterned psychosocial stressors, physiological & behavioral responses, and genomic pathways contribute to the increased risk of HR- breast cancer among Black women. This work provides a basis for exploring potential alternative pathways linking the lived experience of race to the risk of HR- breast cancer, and suggests new avenues for research and public health action.

| [1] |
Anderson WF, Rosenberg PS, Prat A, et al. (2014) How many etiological subtypes of breast cancer: Two, three, four, or more? J Natl Cancer Inst 106: dju165. doi: 10.1093/jnci/dju165

|
| [2] |
Sims AH, Howell A, Howell SJ, et al. (2007) Origins of breast cancer subtypes and therapeutic implications. Nat Clin Pract Oncol 4: 516-525. doi: 10.1038/ncponc0908
|
| [3] | Paquet ER, Hallett MT (2014) Absolute assignment of breast cancer intrinsic molecular subtype. J Natl Cancer Inst 107: 357. |
| [4] |
Perou CM, Sorlie T, Eisen MB, et al. (2000) Molecular portraits of human breast tumours. Nature 406: 747-752. doi: 10.1038/35021093
|
| [5] |
Sorlie T, Tibshirani R, Parker J, et al. (2003) Repeated observation of breast tumor subtypes in independent gene expression data sets. Proc Natl Acad Sci U S A 100: 8418-8423. doi: 10.1073/pnas.0932692100
|
| [6] |
Amend K, Hicks D, Ambrosone CB (2006) Breast cancer in African-American women: differences in tumor biology from European-American women. Cancer Res 66: 8327-8330. doi: 10.1158/0008-5472.CAN-06-1927
|
| [7] | Howlader N, Altekruse SF, Li CI, et al. (2014) US incidence of breast cancer subtypes defined by joint hormone receptor and HER2 Status. J Natl Cancer Inst 106: dju055. |
| [8] |
Newman LA (2014) Breast cancer disparities: high-risk breast cancer and African ancestry. Surg Oncol Clin N Am 23: 579-592. doi: 10.1016/j.soc.2014.03.014
|
| [9] |
Dietze EC, Sistrunk C, Miranda-Carboni G, et al. (2015) Triple-negative breast cancer in African-American women: disparities versus biology. Nat Rev Cancer 15: 248-254. doi: 10.1038/nrc3896
|
| [10] | Gapstur SM, Dupuis J, Gann P, et al. (1996) Hormone receptor status of breast tumors in black, Hispanic, and non-Hispanic white women. An analysis of 13,239 cases. Cancer 77: 1465-1471. |
| [11] |
Hausauer AK, Keegan TH, Chang ET, et al. (2007) Recent breast cancer trends among Asian/Pacific Islander, Hispanic, and African-American women in the US: changes by tumor subtype. Breast Cancer Res 9: R90. doi: 10.1186/bcr1839
|
| [12] |
Joslyn SA (2002) Hormone receptors in breast cancer: racial differences in distribution and survival. Breast Cancer Res Treat 73: 45-59. doi: 10.1023/A:1015220420400
|
| [13] |
Tarone RE, Chu KC (2002) The greater impact of menopause on ER- than ER+ breast cancer incidence: a possible explanation (United States). Cancer Causes Control 13: 7-14. doi: 10.1023/A:1013960609008
|
| [14] |
Dunnwald LK, Rossing MA, Li CI (2007) Hormone receptor status, tumor characteristics, and prognosis: a prospective cohort of breast cancer patients. Breast Cancer Res 9: R6. doi: 10.1186/bcr1639
|
| [15] |
Bauer KR, Brown M, Cress RD, et al. (2007) Descriptive analysis of estrogen receptor (ER)negative, progesterone receptor (PR)-negative, and HER2-negative invasive breast cancer, the so-called triple-negative phenotype - A population-based study from the California Cancer Registry. Cancer 109: 1721-1728. doi: 10.1002/cncr.22618
|
| [16] |
Sineshaw HM, Gaudet M, Ward EM, et al. (2014) Association of race/ethnicity, socioeconomic status, and breast cancer subtypes in the National Cancer Data Base (2010-2011). Breast Cancer Res Treat 145: 753-763. doi: 10.1007/s10549-014-2976-9
|
| [17] | Kohler BA, Sherman RL, Howlader N, et al. (2015) Annual Report to the Nation on the Status of Cancer, 1975-2011, Featuring Incidence of Breast Cancer Subtypes by Race/Ethnicity, Poverty, and State. J Natl Cancer Inst 107: djv048. |
| [18] |
Morris GJ, Naidu S, Topham AK, et al. (2007) Differences in breast carcinoma characteristics in newly diagnosed African-American and Caucasian patients: a single-institution compilation compared with the National Cancer Institute's Surveillance, Epidemiology, and End Results database. Cancer 110: 876-884. doi: 10.1002/cncr.22836
|
| [19] |
Hayanga AJ, Newman LA (2007) Investigating the phenotypes and genotypes of breast cancer in women with African ancestry: The need for more genetic epidemiology. Surg Clin North Am 87: 551-568. doi: 10.1016/j.suc.2007.01.003
|
| [20] |
Newman LA (2015) Disparities in breast cancer and African ancestry: A global perspective. Breast J 21: 133-139. doi: 10.1111/tbj.12369
|
| [21] |
King MC, Marks JH, Mandell JB (2003) Breast and ovarian cancer risks due to inherited mutations in BRCA1 and BRCA2. Science 302: 643-646. doi: 10.1126/science.1088759
|
| [22] |
Churpek JE, Walsh T, Zheng Y, et al. (2015) Inherited predisposition to breast cancer among African American women. Breast Cancer Res Treat 149: 31-39. doi: 10.1007/s10549-014-3195-0
|
| [23] |
Slavich GM, Cole SW (2013) The Emerging Field of Human Social Genomics. Clin Psychol Sci 1: 331-348. doi: 10.1177/2167702613478594
|
| [24] | Geronimus AT (1992) The weathering hypothesis and the health of African-American women and infants: evidence and speculations. Ethn Dis 2: 207-221. |
| [25] | Geronimus AT (1994) The weathering hypothesis and the health of African American women and infants: Implications for reporducitve strategies and policy analysis. In: Sen G, Snow RC, editors. Power and Decision: The Social Control of Reproduction. Cambridge, Mass.: Harvard University Press. |
| [26] | Geronimus AT (2001) Understanding and eliminating racial inequalities in women's health in the United States: the role of the weathering conceptual framework. J Am Med Womens Assoc 56: 133-136, 149-150. |
| [27] | Geronimus AT, Thompson JP (2004) To denigrate, ignore, or disrupt: The health impact of policy-induced breakdown of urban African American communites of support. Du Bois Rev 1: 247-279. |
| [28] |
Geronimus AT, Hicken M, Keene D, et al. (2006) "Weathering" and age patterns of allostatic load scores among blacks and whites in the United States. Am J Public Health 96: 826-833. doi: 10.2105/AJPH.2004.060749
|
| [29] | Geronimus AT, Bound J, Keene D, et al. (2007) Black-white differences in age trajectories of hypertension prevalence among adult women and men, 1999-2002. Ethn Dis 17: 40-48. |
| [30] | Geronimus AT, Hicken MT, Pearson JA, et al. (2010) Do US Black Women Experience Stress-Related Accelerated Biological Aging?: A Novel Theory and First Population-Based Test of Black-White Differences in Telomere Length. Hum Nat 21: 19-38. |
| [31] |
Keene DE, Geronimus AT (2011) Community-based support among African American public housing residents. J Urban Health 88: 41-53. doi: 10.1007/s11524-010-9511-z
|
| [32] |
Geronimus AT (2013) Deep integration: Letting the epigenome out of the bottle without losing sight of the structural origins of population Health. Am J Public Health 103: S56-S63. doi: 10.2105/AJPH.2013.301380
|
| [33] |
Williams DR, Mohammed SA, Shields AE (2016) Understanding and effectively addressing breast cancer in African American women: Unpacking the social context. Cancer 122: 2138-2149. doi: 10.1002/cncr.29935
|
| [34] |
Trivers K, Lund M, Porter P, et al. (2009) The epidemiology of triple-negative breast cancer, including race. Cancer Causes Control 20: 1071-1082. doi: 10.1007/s10552-009-9331-1
|
| [35] |
Kwan ML, Kushi LH, Weltzien E, et al. (2009) Epidemiology of breast cancer subtypes in two prospective cohort studies of breast cancer survivors. Breast Cancer Res 11: R31. doi: 10.1186/bcr2261
|
| [36] |
Millikan RC, Newman B, Tse CK, et al. (2008) Epidemiology of basal-like breast cancer. Breast Cancer Res Treat 109: 123-139. doi: 10.1007/s10549-007-9632-6
|
| [37] |
Parise CA, Bauer KR, Brown MM, et al. (2009) Breast cancer subtypes as defined by the estrogen receptor (ER), progesterone receptor (PR), and the human epidermal growth factor receptor 2 (HER2) among women with invasive breast cancer in California, 1999-2004. Breast J 15: 593-602. doi: 10.1111/j.1524-4741.2009.00822.x
|
| [38] |
Carey LA, Perou CM, Livasy CA, et al. (2006) Race, breast cancer subtypes, and survival in the Carolina Breast Cancer Study. JAMA 295: 2492-2502. doi: 10.1001/jama.295.21.2492
|
| [39] |
Setiawan VW, Monroe KR, Wilkens LR, et al. (2009) Breast cancer risk factors defined by estrogen and progesterone receptor status: the multiethnic cohort study. Am J Epidemiol 169: 1251-1259. doi: 10.1093/aje/kwp036
|
| [40] |
Gehlert S, Sohmer D, Sacks T, et al. (2008) Targeting health disparities: a model linking upstream determinants to downstream interventions. Health Aff (Millwood) 27: 339-349. doi: 10.1377/hlthaff.27.2.339
|
| [41] |
Warnecke RB, Oh A, Breen N, et al. (2008) Approaching health disparities from a population perspective: the National Institutes of Health Centers for population health and health disparities. Am J Public Health 98: 1608-1615. doi: 10.2105/AJPH.2006.102525
|
| [42] |
Pearlin LI, Schieman S, Fazio EM, et al. (2005) Stress, health, and the life course: Some conceptual perspectives. J Health Soc Behav 46: 205-219. doi: 10.1177/002214650504600206
|
| [43] |
Hill TD, Ross CE, Angel RJ (2005) Neighborhood disorder, psychophysiological distress, and health. J Health Soc Behav 46: 170-186. doi: 10.1177/002214650504600204
|
| [44] |
Wheaton B (1985) Models for the stress-buffering functions of coping resources. J Health Soc Behav 26: 352-364. doi: 10.2307/2136658
|
| [45] |
Petticrew M, Fraser JM, Regan MF (1999) Adverse life-events and risk of breast cancer: A meta-analysis. Br J Health Psychol 4: 1-17. doi: 10.1348/135910799168434
|
| [46] |
Chida Y, Hamer M, Wardle J, et al. (2008) Do stress-related psychosocial factors contribute to cancer incidence and survival? Nat Clin Pract Oncol 5: 466-475. doi: 10.1038/ncponc1134
|
| [47] |
Holmes TH, Rahe RH (1967) The social readjustment rating scale. Psychosom Med 11: 213-218. doi: 10.1016/0022-3999(67)90010-4
|
| [48] |
Brown JS, Meadows SO, Elder GH, Jr. (2007) Race-ethnic inequality and psychological distress: Depressive symptoms from adolescence to young adulthood. Dev Psychol 43: 1295-1311. doi: 10.1037/0012-1649.43.6.1295
|
| [49] |
Geronimus AT, Pearson JA, Linnenbringer E, et al. (2015) Race-ethnicity, poverty, urban stressors, and telomere length in a detroit community-based sample. J Health Soc Behav 56: 199-224. doi: 10.1177/0022146515582100
|
| [50] | Cheang A, Cooper CL (1985) Psychosocial factors in breast cancer. Stress Med 1: 11-24. |
| [51] |
Sorlie T (2004) Molecular portraits of breast cancer: tumour subtypes as distinct disease entities. Eur J Cancer 40: 2667-2675. doi: 10.1016/j.ejca.2004.08.021
|
| [52] |
Chrousos GP, Torpy DJ, Gold PW (1998) Interactions between the hypothalamic-pituitary-adrenal axis and the female reproductive system: clinical implications. Ann Intern Med 129: 229-240. doi: 10.7326/0003-4819-129-3-199808010-00012
|
| [53] |
Spiegel D, Butler LD, Giese-Davis J, et al. (2007) Effects of supportive-expressive group therapy on survival of patients with metastatic breast cancer: a randomized prospective trial. Cancer 110: 1130-1138. doi: 10.1002/cncr.22890
|
| [54] |
Michael YL, Carlson NE, Chlebowski RT, et al. (2009) Influence of stressors on breast cancer incidence in the Women's Health Initiative. Health Psychol 28: 137-146. doi: 10.1037/a0012982
|
| [55] |
Melhem-Bertrandt A, Conzen S (2010) The relationship between psychosocial stressors and breast cancer biology. Current Breast Cancer Reports 2: 130-137. doi: 10.1007/s12609-010-0021-5
|
| [56] |
Cacioppo JT, Hawkley LC (2003) Social isolation and health, with an emphasis on underlying mechanisms. Perspect Biol Med 46: S39-52. doi: 10.1353/pbm.2003.0049
|
| [57] | McClintock MK, Conzen SD, Gehlert S, et al. (2005) Mammary cancer and social interactions: identifying multiple environments that regulate gene expression throughout the life span. J Gerontol B Psychol Sci Soc Sci 60 1: 32-41. |
| [58] |
Williams JB, Pang D, Delgado B, et al. (2009) A model of gene-environment interaction reveals altered mammary gland gene expression and increased tumor growth following social isolation. Cancer Prev Res 2: 850-861. doi: 10.1158/1940-6207.CAPR-08-0238
|
| [59] |
Hasen NS, O'Leary KA, Auger AP, et al. (2010) Social isolation reduces mammary development, tumor incidence, and expression of epigenetic regulators in wild-type and p53-heterozygotic mice. Cancer Prev Res (Phila Pa) 3: 620-629. doi: 10.1158/1940-6207.CAPR-09-0225
|
| [60] |
Williams DR, Neighbors HW, Jackson JS (2003) Racial/ethnic discrimination and health: findings from community studies. Am J Public Health 93: 200-208. doi: 10.2105/AJPH.93.2.200
|
| [61] |
Taylor TR, Williams CD, Makambi KH, et al. (2007) Racial discrimination and breast cancer incidence in US black women - the black women's health study. Am J Epidemiol 166: 46-54. doi: 10.1093/aje/kwm056
|
| [62] |
Krieger N, Jahn JL, Waterman PD (2017) Jim Crow and estrogen-receptor-negative breast cancer: US-born black and white non-Hispanic women, 1992-2012. Cancer Causes Control 28: 49-59. doi: 10.1007/s10552-016-0834-2
|
| [63] |
Williams DR, Collins C (2001) Racial residential segregation: a fundamental cause of racial disparities in health. Public Health Rep 116: 404-416. doi: 10.1016/S0033-3549(04)50068-7
|
| [64] |
Schulz A, Williams DR, Israel BA, et al. (2002) Racial and spatial relations as fundamental determinants of health in Detroit. Milbank Q 80: 677-707. doi: 10.1111/1468-0009.00028
|
| [65] | Massey DS, Denton NA (1993) American Apartheid: Segregation and the Making of the Underclass. Cambridge, Mass.: Harvard University Press. |
| [66] | Barrett RE, Cho YI, Weaver KE, et al. (2008) Neighborhood change and distant metastasis at diagnosis of breast cancer. Ann Epidemiol 18: 43-47. |
| [67] |
Sampson RJ, Morenoff JD, Earls F (1999) Beyond social capital: Spatial dynamics of collective efficacy for children. Am Sociol Rev 64: 633-660. doi: 10.2307/2657367
|
| [68] |
Warner E, Gomez S (2010) Impact of Neighborhood Racial Composition and Metropolitan Residential Segregation on Disparities in Breast Cancer Stage at Diagnosis and Survival Between Black and White Women in California. J Community Health 35: 398-408. doi: 10.1007/s10900-010-9265-2
|
| [69] | Linnenbringer E (2014) Social constructions, biological implications: A structural examination of racial disparities in breast cancer subtype. Ann Arbor, MI: University of Michigan. |
| [70] | Linnenbringer E, Geronimus AT (2014) Neighborhood SES, racial concentration, and hormone receptor status among california women diagnosed with breast cancer population association of america annual meeting. Boston, MA. |
| [71] | Sampson RJ, Morenoff JD, Gannon-Rowley T (2002) Assessing "neighborhood effects": Social processes and new directions in research. Ann RevSociol 28: 443-478. |
| [72] |
Bernard P, Charafeddine R, Frohlich KL, et al. (2007) Health inequalities and place: A theoretical conception of neighbourhood. Soc Sci Med 65: 1839-1852. doi: 10.1016/j.socscimed.2007.05.037
|
| [73] |
Cummins S, Curtis S, Diez-Roux AV, et al. (2007) Understanding and representing 'place' in health research: A relational approach. Soc Sci Med 65: 1825-1838. doi: 10.1016/j.socscimed.2007.05.036
|
| [74] |
Macintyre S, Ellaway A, Cummins S (2002) Place effects on health: how can we conceptualise, operationalise and measure them? Soc Sci Med 55: 125-139. doi: 10.1016/S0277-9536(01)00214-3
|
| [75] |
Pickett KE, Wilkinson RG (2008) People like us: ethnic group density effects on health. Ethn Health 13: 321-334. doi: 10.1080/13557850701882928
|
| [76] | Becares L, Nazroo J, Stafford M (2009) The buffering effects of ethnic density on experienced racism and health. Health Place 15: 670-678. |
| [77] | Bécares L, Shaw R, Nazroo J, et al. (2012) Ethnic density effects on physical morbidity, mortality, and health behaviors: A systematic review of the literature. Am J Public Health 102: e33-e66. |
| [78] |
Keene D, Bader M, Ailshire J (2013) Length of residence and social integration: The contingent effects of neighborhood poverty. Health Place 21: 171-178. doi: 10.1016/j.healthplace.2013.02.002
|
| [79] | Small ML (2004) Villa Victoria: The transformation of coail capital in a Boston Barrio. Chicago: University of Chicago Press. |
| [80] |
Tach LM (2009) More than bricks and mortar: Neighborhood frames, social processes, and the mixed-income redevelopment of a public housing project. City Commun 8: 269-299. doi: 10.1111/j.1540-6040.2009.01289.x
|
| [81] |
Pearlin LI (1989) The sociological study of stress. J Health Soc Behav 30: 241-256. doi: 10.2307/2136956
|
| [82] |
Taylor SE, Repetti RL, Seeman T (1997) Health psychology: What is an unhealthy environment and how does it get under the skin? Ann Rev Psychol 48: 411-447. doi: 10.1146/annurev.psych.48.1.411
|
| [83] | Massey DS (2004) Segregation and stratification: A biosocial perspective. Du Bois Review 1: 7-25. |
| [84] | Jackson JS, Knight KM (2006) Race and self-regulatory health behaviors: The role of the stress response and the HPA axis in physical and mental health disparities. In: Schaie KW, Carstensen LL, editors. Social Structures, Aging, and Self-Regulation in the Elderly. New York: Springer: 189-207. |
| [85] | Cole SW, Hawkley LC, Arevalo JM, et al. (2007) Social regulation of gene expression in human leukocytes. Genome Biol 8(9): R189. |
| [86] |
Traustadóttir T, Bosch PR, Matt KS (2005) The HPA axis response to stress in women: effects of aging and fitness. Psychoneuroendocrinology 30: 392-402. doi: 10.1016/j.psyneuen.2004.11.002
|
| [87] |
McEwen BS (1998) Protective and damaging effects of stress mediators. N Engl J Med 338: 171-179. doi: 10.1056/NEJM199801153380307
|
| [88] |
McEwen BS, Wingfield JC (2003) The concept of allostasis in biology and biomedicine. Horm Behav 43: 2-15. doi: 10.1016/S0018-506X(02)00024-7
|
| [89] |
Parente V, Hale L, Palermo T (2013) Association between breast cancer and allostatic load by race: National Health and Nutrition Examination Survey 1999-2008. Psycho-Oncology 22: 621-628. doi: 10.1002/pon.3044
|
| [90] |
Morland K, Wing S, Diez Roux A, et al. (2002) Neighborhood characteristics associated with the location of food stores and food service places. Am J Prev Med 22: 23-29. doi: 10.1016/S0749-3797(01)00403-2
|
| [91] |
Moore LV, Diez Roux AV (2006) Associations of neighborhood characteristics with the location and type of food stores. Am J Public Health 96: 325-331. doi: 10.2105/AJPH.2004.058040
|
| [92] | Zenk SN, Schulz AJ, Israel BA, et al. (2006) Fruit and vegetable access differs by community racial composition and socioeconomic position in Detroit, Michigan. Ethn Dis 16: 275-280. |
| [93] | Baker EA, Schootman M, Barnidge E, et al. (2006) The role of race and poverty in access to foods that enable individuals to adhere to dietary guidelines. Prev Chronic Dis 3: A76. |
| [94] |
Jackson JS, Knight KM, Rafferty JA (2010) Race and unhealthy behaviors: chronic stress, the HPA axis, and physical and mental health disparities over the life course. Am J Public Health 100: 933-939. doi: 10.2105/AJPH.2008.143446
|
| [95] |
Zhang SM, Hankinson SE, Hunter DJ, et al. (2005) Folate intake and risk of breast cancer characterized by hormone receptor status. Cancer Epidemiol Biomarkers Prev 14: 2004-2008. doi: 10.1158/1055-9965.EPI-05-0083
|
| [96] |
Boggs DA, Palmer JR, Wise LA, et al. (2010) Fruit and vegetable intake in relation to risk of breast cancer in the black women's health study. Am J Epidemiol 172: 1268-1279. doi: 10.1093/aje/kwq293
|
| [97] |
Esteller M (2008) Epigenetics in cancer. N Engl J Med 358: 1148-1159. doi: 10.1056/NEJMra072067
|
| [98] |
Gronbaek K, Hother C, Jones PA (2007) Epigenetic changes in cancer. Apmis 115: 1039-1059. doi: 10.1111/j.1600-0463.2007.apm_636.xml.x
|
| [99] | Ahuja N, Li Q, Mohan AL, et al. (1998) Aging and DNA methylation in colorectal mucosa and cancer. Cancer Res 58: 5489-5494. |
| [100] | Issa JP (2000) CpG-island methylation in aging and cancer. Curr Top Microbiol Immunol 249: 101-118. |
| [101] |
Szyf M, McGowan P, Meaney MJ (2008) The social environment and the epigenome. Environ Mol Mutagen 49: 46-60. doi: 10.1002/em.20357
|
| [102] |
Fraga MF, Ballestar E, Paz MF, et al. (2005) Epigenetic differences arise during the lifetime of monozygotic twins. Proc Natl Acad Sci U S A 102: 10604-10609. doi: 10.1073/pnas.0500398102
|
| [103] |
Jovanovic J, Rønneberg JA, Tost J, et al. (2010) The epigenetics of breast cancer. Mol Oncol 4: 242-254. doi: 10.1016/j.molonc.2010.04.002
|
| [104] | Weigel RJ, deConinck EC (1993) Transcriptional control of estrogen receptor in estrogen receptor-negative breast carcinoma. Cancer Res 53: 3472-3474. |
| [105] | Ferguson AT, Lapidus RG, Baylin SB, et al. (1995) Demethylation of the estrogen receptor gene in estrogen receptor-negative breast cancer cells can reactivate estrogen receptor gene expression. Cancer Res 55: 2279-2283. |
| [106] | Wei M, Xu J, Dignam J, et al. (2007) Estrogen receptor alpha, BRCA1, and FANCF promoter methylation occur in distinct subsets of sporadic breast cancers. Breast Cancer Res Treat 111: 113-120. |
| [107] |
Gaudet MM, Campan M, Figueroa JD, et al. (2009) DNA hypermethylation of ESR1 and PGR in breast cancer: pathologic and epidemiologic associations. Cancer Epidemiol Biomarkers Prev 18: 3036-3043. doi: 10.1158/1055-9965.EPI-09-0678
|
| [108] |
Feng W, Shen L, Wen S, et al. (2007) Correlation between CpG methylation profiles and hormone receptor status in breast cancers. Breast Cancer Res 9: R57. doi: 10.1186/bcr1762
|
| [109] |
Widschwendter M, Siegmund KD, Muller HM, et al. (2004) Association of breast cancer DNA methylation profiles with hormone receptor status and response to tamoxifen. Cancer Res 64: 3807-3813. doi: 10.1158/0008-5472.CAN-03-3852
|
| [110] |
Christensen BC, Kelsey KT, Zheng S, et al. (2010) Breast Cancer DNA Methylation Profiles Are Associated with Tumor Size and Alcohol and Folate Intake. PLoS Genet 6: e1001043. doi: 10.1371/journal.pgen.1001043
|
| [111] |
Cole SW (2010) Elevating the perspective on human stress genomics. Psychoneuroendocrinology 35: 955-962. doi: 10.1016/j.psyneuen.2010.06.008
|
Figures(1) / Tables(1)

Erin Linnenbringer, Sarah Gehlert, Arline T. Geronimus. Black-White Disparities in Breast Cancer Subtype: The Intersection of Socially Patterned Stress and Genetic Expression[J]. AIMS Public Health, 2017, 4(5): 526-556. doi: 10.3934/publichealth.2017.5.526







 DownLoad:
DownLoad: