Citation: Joanne L. Hopper, Natasha Begum, Laura Smith, Thomas A. Hughes. The role of PSMD9 in human disease: future clinical and therapeutic implications[J]. AIMS Molecular Science, 2015, 2(4): 476-484. doi: 10.3934/molsci.2015.4.476

| [1] |
Uhlen M, Oksvold P, Fagerberg L, et al. (2010) Towards a knowledge-based Human Protein Atlas. Nat Biotechnol 28: 1248-1250. doi: 10.1038/nbt1210-1248

|
| [2] |
Watanabe TK, Saito A, Suzuki M, et al. (1998) cDNA cloning and characterization of a human proteasomal modulator subunit, p27 (PSMD9). Genomics 50: 241-250. doi: 10.1006/geno.1998.5301
|
| [3] |
Stanojevic V, Yao KM, Thomas MK (2005) The coactivator Bridge-1 increases transcriptional activation by pancreas duodenum homeobox-1 (PDX-1). Mol Cell Endocrinol 237: 67-74. doi: 10.1016/j.mce.2005.03.003
|
| [4] |
Thomas MK, Yao KM, Tenser MS, et al. (1999) Bridge-1, a novel PDZ-domain coactivator of E2A-mediated regulation of insulin gene transcription. Mol Cell Biol 19: 8492-8504. doi: 10.1128/MCB.19.12.8492
|
| [5] |
Dalton WS (2004) The proteasome. Semin Oncol 31: 3-9. doi: 10.1053/S0093-7754(04)00498-1
|
| [6] |
Sahu I, Sangith N, Ramteke M, et al. (2014) A novel role for the proteasomal chaperone PSMD9 and hnRNPA1 in enhancing IkappaBalpha degradation and NF-kappaB activation - functional relevance of predicted PDZ domain-motif interaction. FEBS J 281: 2688-2709. doi: 10.1111/febs.12814
|
| [7] |
Banz-Jansen C, Munchow B, Diedrich K, et al. (2011) Bridge-1 is expressed in human breast carcinomas: silencing of Bridge-1 decreases Smad2, Smad3 and Smad4 expression in MCF-7 cells, a human breast cancer cell line. Arch Gynecol Obstet 284: 1543-1549. doi: 10.1007/s00404-011-1875-0
|
| [8] |
de Kretser DM, O'Hehir RE, Hardy CL, et al. (2012) The roles of activin A and its binding protein, follistatin, in inflammation and tissue repair. Mol Cell Endocrinol 359: 101-106. doi: 10.1016/j.mce.2011.10.009
|
| [9] |
Hashimoto O, Funaba M (2011) Activin in glucose metabolism. Vitam Horm 85: 217-234. doi: 10.1016/B978-0-12-385961-7.00011-1
|
| [10] |
Deli A, Kreidl E, Santifaller S, et al. (2008) Activins and activin antagonists in hepatocellular carcinoma. World J Gastroenterol 14: 1699-1709. doi: 10.3748/wjg.14.1699
|
| [11] |
Ottley E, Gold E (2012) Insensitivity to the growth inhibitory effects of activin A: an acquired capability in prostate cancer progression. Cytokine Growth Factor Rev 23: 119-125. doi: 10.1016/j.cytogfr.2012.04.004
|
| [12] |
Loomans HA, Andl CD (2014) Intertwining of Activin A and TGFbeta Signaling: Dual Roles in Cancer Progression and Cancer Cell Invasion. Cancers (Basel) 7: 70-91. doi: 10.3390/cancers7010070
|
| [13] |
Langlands FE, Dodwell D, Hanby AM, et al. (2014) PSMD9 expression predicts radiotherapy response in breast cancer. Mol Cancer 13: 73. doi: 10.1186/1476-4598-13-73
|
| [14] |
Alsner J, Sorensen SB, Overgaard J (2001) TP53 mutation is related to poor prognosis after radiotherapy, but not surgery, in squamous cell carcinoma of the head and neck. Radiother Oncol 59: 179-185. doi: 10.1016/S0167-8140(01)00301-2
|
| [15] | Abdel Raheem AM, Hameed DA, ElGanainy EO, et al. (2011) Can Bcl-XL expression predict the radio sensitivity of bilharzial-related squamous bladder carcinoma? A prospective comparative study. BMC Cancer 11: 16. |
| [16] |
Asanuma K, Moriai R, Yajima T, et al. (2000) Survivin as a radioresistance factor in pancreatic cancer. Jpn J Cancer Res 91: 1204-1209. doi: 10.1111/j.1349-7006.2000.tb00906.x
|
| [17] |
Crawford LJ, Walker B, Irvine AE (2011) Proteasome inhibitors in cancer therapy. J Cell Commun Signal 5: 101-110. doi: 10.1007/s12079-011-0121-7
|
| [18] |
Hanis CL, Boerwinkle E, Chakraborty R, et al. (1996) A genome-wide search for human non-insulin-dependent (type 2) diabetes genes reveals a major susceptibility locus on chromosome 2. Nat Genet 13: 161-166. doi: 10.1038/ng0696-161
|
| [19] |
Mahtani MM, Widen E, Lehto M, et al. (1996) Mapping of a gene for type 2 diabetes associated with an insulin secretion defect by a genome scan in Finnish families. Nat Genet 14: 90-94. doi: 10.1038/ng0996-90
|
| [20] |
Gragnoli C (2010) PSMD9 gene in the NIDDM2 locus is linked to type 2 diabetes in Italians. J Cell Physiol 222: 265-267. doi: 10.1002/jcp.21954
|
| [21] |
Gragnoli C, Cronsell J (2007) PSMD9 gene variants within NIDDM2 may rarely contribute to type 2 diabetes. J Cell Physiol 212: 568-571. doi: 10.1002/jcp.21127
|
| [22] | Gragnoli C (2010) PSMD9 is linked to MODY3. J Cell Physiol 223: 1-5. |
| [23] |
Gragnoli C (2011) PSMD9 is linked to type 2 diabetes neuropathy. J Diabetes Complications 25: 329-331. doi: 10.1016/j.jdiacomp.2011.06.003
|
| [24] |
Gragnoli C (2012) Proteasome modulator 9 is linked to microvascular pathology of T2D. J Cell Physiol 227: 3116-3118. doi: 10.1002/jcp.23063
|
| [25] |
Gragnoli C (2011) Proteasome modulator 9 and macrovascular pathology of T2D. Cardiovasc Diabetol 10: 32. doi: 10.1186/1475-2840-10-32
|
| [26] |
Gragnoli C (2011) Proteasome modulator 9 SNPs are linked to hypertension in type 2 diabetes families. Cardiovasc Diabetol 10: 77. doi: 10.1186/1475-2840-10-77
|
| [27] |
Lee JH, Volinic JL, Banz C, et al. (2005) Interactions with p300 enhance transcriptional activation by the PDZ-domain coactivator Bridge-1. J Endocrinol 187: 283-292. doi: 10.1677/joe.1.06305
|
| [28] |
Thomas MK, Tsang SW, Yeung ML, et al. (2009) The roles of the PDZ-containing proteins bridge-1 and PDZD2 in the regulation of insulin production and pancreatic beta-cell mass. Curr Protein Pept Sci 10: 30-36. doi: 10.2174/138920309787315248
|
| [29] |
Volinic JL, Lee JH, Eto K, et al. (2006) Overexpression of the coactivator bridge-1 results in insulin deficiency and diabetes. Mol Endocrinol 20: 167-182. doi: 10.1210/me.2005-0127
|
| [30] |
Donath MY (2014) Targeting inflammation in the treatment of type 2 diabetes: time to start. Nat Rev Drug Discov 13: 465-476. doi: 10.1038/nrd4275
|
| [31] |
Liu H, Yu S, Xu W, et al. (2012) Enhancement of 26S proteasome functionality connects oxidative stress and vascular endothelial inflammatory response in diabetes mellitus. Arterioscler Thromb Vasc Biol 32: 2131-2140. doi: 10.1161/ATVBAHA.112.253385
|
| [32] |
Sullivan PF, Neale MC, Kendler KS (2000) Genetic epidemiology of major depression: review and meta-analysis. Am J Psychiatry 157: 1552-1562. doi: 10.1176/appi.ajp.157.10.1552
|
| [33] |
Millier A, Schmidt U, Angermeyer MC, et al. (2014) Humanistic burden in schizophrenia: a literature review. J Psychiatr Res 54: 85-93. doi: 10.1016/j.jpsychires.2014.03.021
|
| [34] |
Cardno AG, Marshall EJ, Coid B, et al. (1999) Heritability estimates for psychotic disorders: the Maudsley twin psychosis series. Arch Gen Psychiatry 56: 162-168. doi: 10.1001/archpsyc.56.2.162
|
| [35] |
Mezuk B, Eaton WW, Albrecht S, et al. (2008) Depression and type 2 diabetes over the lifespan: a meta-analysis. Diabetes Care 31: 2383-2390. doi: 10.2337/dc08-0985
|
| [36] | Pan A, Lucas M, Sun Q, et al. (2010) Bidirectional association between depression and type 2 diabetes mellitus in women. Arch Intern Med 170: 1884-1891. |
| [37] |
Gragnoli C (2014) Proteasome modulator 9 gene SNPs, responsible for anti-depressant response, are in linkage with generalized anxiety disorder. J Cell Physiol 229: 1157-1159. doi: 10.1002/jcp.24581
|
| [38] |
Gragnoli C (2012) Proteasome modulator 9 and depression in type 2 diabetes. Curr Med Chem 19: 5178-5180. doi: 10.2174/092986712803530593
|
| [39] |
Wong ML, Dong C, Maestre-Mesa J, et al. (2008) Polymorphisms in inflammation-related genes are associated with susceptibility to major depression and antidepressant response. Mol Psychiatry 13: 800-812. doi: 10.1038/mp.2008.59
|
| [40] |
Wong ML, Dong C, Andreev V, et al. (2012) Prediction of susceptibility to major depression by a model of interactions of multiple functional genetic variants and environmental factors. Mol Psychiatry 17: 624-633. doi: 10.1038/mp.2012.13
|
| [41] |
Lee YH, Kim JH, Song GG (2013) Pathway analysis of a genome-wide association study in schizophrenia. Gene 525: 107-115. doi: 10.1016/j.gene.2013.04.014
|
| [42] |
Haase J, Brown E (2015) Integrating the monoamine, neurotrophin and cytokine hypotheses of depression--a central role for the serotonin transporter? Pharmacol Ther 147: 1-11. doi: 10.1016/j.pharmthera.2014.10.002
|
| [43] |
Furtado M, Katzman MA (2015) Examining the role of neuroinflammation in major depression. Psychiatry Res 229: 27-36. doi: 10.1016/j.psychres.2015.06.009
|
| [44] | Uzbekov M, Maxinova N (2015) Biochemical Bases of Monoamine and Hormonal Interactions in Pathogenesis of Anxious Depression: a Hypothesis. European Psychiatry 30: 542. |
| [45] |
Najjar S, Pearlman DM (2015) Neuroinflammation and white matter pathology in schizophrenia: systematic review. Schizophr Res 161: 102-112. doi: 10.1016/j.schres.2014.04.041
|
| [46] |
Talbot K, Eidem WL, Tinsley CL, et al. (2004) Dysbindin-1 is reduced in intrinsic, glutamatergic terminals of the hippocampal formation in schizophrenia. J Clin Invest 113: 1353-1363. doi: 10.1172/JCI200420425
|
| [47] | Weickert CS, Rothmond DA, Hyde TM, et al. (2008) Reduced DTNBP1 (dysbindin-1) mRNA in the hippocampal formation of schizophrenia patients. Schizophr Res 98: 105-110. |
| [48] |
Saggu S, Cannon TD, Jentsch JD, et al. (2013) Potential molecular mechanisms for decreased synaptic glutamate release in dysbindin-1 mutant mice. Schizophr Res 146: 254-263. doi: 10.1016/j.schres.2013.01.037
|
| [49] |
Tang J, LeGros RP, Louneva N, et al. (2009) Dysbindin-1 in dorsolateral prefrontal cortex of schizophrenia cases is reduced in an isoform-specific manner unrelated to dysbindin-1 mRNA expression. Hum Mol Genet 18: 3851-3863. doi: 10.1093/hmg/ddp329
|
| [50] |
Han MH, Hu Z, Chen CY, et al. (2014) Dysbindin-associated proteome in the p2 synaptosome fraction of mouse brain. J Proteome Res 13: 4567-4580. doi: 10.1021/pr500656z
|
| [51] |
Banz C, Munchow B, Diedrich K (2010) Bridge-1 is expressed in human granulosa cells and is involved in the activin A signaling cascade. Fertil Steril 93: 1349-1352. doi: 10.1016/j.fertnstert.2009.07.1675
|
| [52] |
Barber TM, Franks S (2012) The link between polycystic ovary syndrome and both Type 1 and Type 2 diabetes mellitus: what do we know today? Womens Health (Lond Engl) 8: 147-154. doi: 10.2217/whe.11.94
|
| [53] |
Ehrmann DA (2005) Polycystic ovary syndrome. N Engl J Med 352: 1223-1236. doi: 10.1056/NEJMra041536
|
| [54] | Marti MJ, Tolosa E, Campdelacreu J (2003) Clinical overview of the synucleinopathies. Mov Disord 18 Suppl 6: S21-27. |
| [55] |
Stefanova N, Klimaschewski L, Poewe W, et al. (2001) Glial cell death induced by overexpression of alpha-synuclein. J Neurosci Res 65: 432-438. doi: 10.1002/jnr.1171
|
| [56] |
Xilouri M, Brekk OR, Stefanis L (2013) alpha-Synuclein and protein degradation systems: a reciprocal relationship. Mol Neurobiol 47: 537-551. doi: 10.1007/s12035-012-8341-2
|
| [57] | Stefanis L (2012) alpha-Synuclein in Parkinson's disease. Cold Spring Harb Perspect Med 2: a009399. |
| [58] | Vartiainen S, Pehkonen P, Lakso M, et al. (2006) Identification of gene expression changes in transgenic C. elegans overexpressing human alpha-synuclein. Neurobiol Dis 22: 477-486. |
| [59] |
Rideout HJ, Dietrich P, Wang Q, et al. (2004) alpha-synuclein is required for the fibrillar nature of ubiquitinated inclusions induced by proteasomal inhibition in primary neurons. J Biol Chem 279: 46915-46920. doi: 10.1074/jbc.M405146200
|
| [60] | Pierre S-R, Vernace V, Wang Z, et al. (2009) Mechanisms Linking the Ubiquitin/Proteasome Pathway and Chaperones. In: Richter-Landsberg C, editor. Heat Shock Proteins in Neural Cells: Springer New York. |
| [61] |
Bedford L, Hay D, Devoy A, et al. (2008) Depletion of 26S proteasomes in mouse brain neurons causes neurodegeneration and Lewy-like inclusions resembling human pale bodies. J Neurosci 28: 8189-8198. doi: 10.1523/JNEUROSCI.2218-08.2008
|
| [62] |
Sangith N, Srinivasaraghavan K, Sahu I, et al. (2014) Discovery of novel interacting partners of PSMD9, a proteasomal chaperone: Role of an Atypical and versatile PDZ-domain motif interaction and identification of putative functional modules. FEBS Open Bio 4: 571-583. doi: 10.1016/j.fob.2014.05.005
|
| [63] |
Geng X, Lou H, Wang J, et al. (2011) alpha-Synuclein binds the K(ATP) channel at insulin-secretory granules and inhibits insulin secretion. Am J Physiol Endocrinol Metab 300: E276-286. doi: 10.1152/ajpendo.00262.2010
|
| [64] |
Steneberg P, Bernardo L, Edfalk S, et al. (2013) The type 2 diabetes-associated gene ide is required for insulin secretion and suppression of alpha-synuclein levels in beta-cells. Diabetes 62: 2004-2014. doi: 10.2337/db12-1045
|
| [65] |
Hu G, Jousilahti P, Bidel S, et al. (2007) Type 2 diabetes and the risk of Parkinson's disease. Diabetes Care 30: 842-847. doi: 10.2337/dc06-2011
|
| [66] |
Cereda E, Barichella M, Cassani E, et al. (2012) Clinical features of Parkinson disease when onset of diabetes came first: A case-control study. Neurology 78: 1507-1511. doi: 10.1212/WNL.0b013e3182553cc9
|
| [67] |
Cereda E, Barichella M, Pedrolli C, et al. (2011) Diabetes and risk of Parkinson's disease: a systematic review and meta-analysis. Diabetes Care 34: 2614-2623. doi: 10.2337/dc11-1584
|
| [68] |
Sandyk R (1993) The relationship between diabetes mellitus and Parkinson's disease. Int J Neurosci 69: 125-130. doi: 10.3109/00207459309003322
|
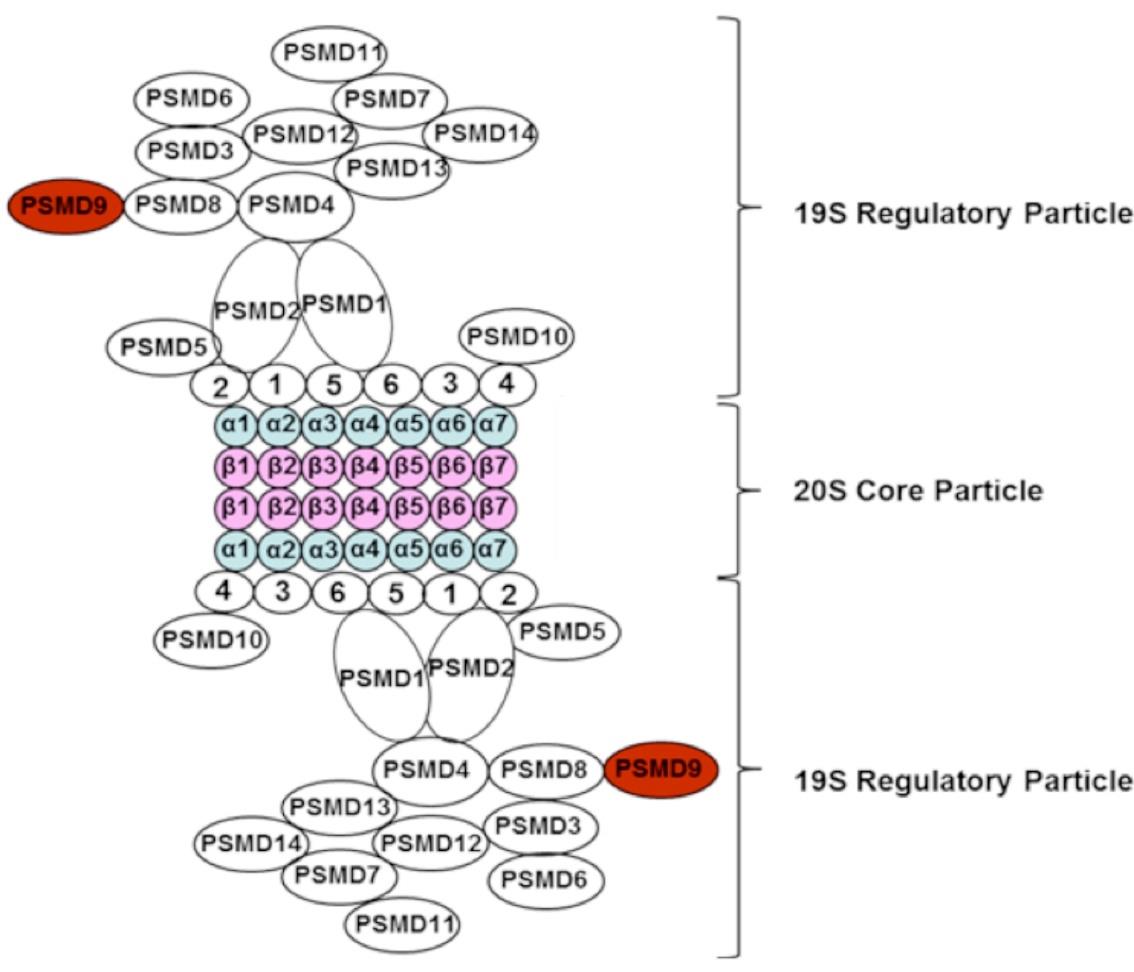
Figures(1)

Joanne L. Hopper, Natasha Begum, Laura Smith, Thomas A. Hughes. The role of PSMD9 in human disease: future clinical and therapeutic implications[J]. AIMS Molecular Science, 2015, 2(4): 476-484. doi: 10.3934/molsci.2015.4.476







 DownLoad:
DownLoad: