Citation: Tsuyoshi Inoshita, Yuzuru Imai. Regulation of vesicular trafficking by Parkinson's disease-associated genes[J]. AIMS Molecular Science, 2015, 2(4): 461-475. doi: 10.3934/molsci.2015.4.461

| [1] |
Sudhof TC (2004) The synaptic vesicle cycle. Annu Rev Neurosci 27: 509-547. doi: 10.1146/annurev.neuro.26.041002.131412

|
| [2] |
Schweizer FE, Ryan TA (2006) The synaptic vesicle: cycle of exocytosis and endocytosis. Curr Opin Neurobiol 16: 298-304. doi: 10.1016/j.conb.2006.05.006
|
| [3] |
Dittman J, Ryan TA (2009) Molecular circuitry of endocytosis at nerve terminals. Annu Rev Cell Dev Biol 25: 133-160. doi: 10.1146/annurev.cellbio.042308.113302
|
| [4] |
Kuromi H, Kidokoro Y (1998) Two distinct pools of synaptic vesicles in single presynaptic boutons in a temperature-sensitive Drosophila mutant, shibire. Neuron 20: 917-925. doi: 10.1016/S0896-6273(00)80473-0
|
| [5] |
Richards DA, Guatimosim C, Betz WJ (2000) Two endocytic recycling routes selectively fill two vesicle pools in frog motor nerve terminals. Neuron 27: 551-559. doi: 10.1016/S0896-6273(00)00065-9
|
| [6] |
Mohrmann R, de Wit H, Connell E, et al. (2013) Synaptotagmin interaction with SNAP-25 governs vesicle docking, priming, and fusion triggering. J Neurosci 33: 14417-14430. doi: 10.1523/JNEUROSCI.1236-13.2013
|
| [7] |
Verstreken P, Ly CV, Venken KJT, et al. (2005) Synaptic Mitochondria Are Critical for Mobilization of Reserve Pool Vesicles at Drosophila Neuromuscular Junctions. Neuron 47: 365-378. doi: 10.1016/j.neuron.2005.06.018
|
| [8] |
Pelassa I, Zhao C, Pasche M, et al. (2014) Synaptic vesicles are . Front Mol Neurosci 7: 91-. doi: 10.3389/fnmol.2014.00091
|
| [9] | Benmerah A, Bayrou M, Cerf-Bensussan N, et al. (1999) Inhibition of clathrin-coated pit assembly by an Eps15 mutant. J Cell Sci 112 (Pt 9): 1303-1311. |
| [10] |
Watanabe S, Trimbuch T, Camacho-Perez M, et al. (2014) Clathrin regenerates synaptic vesicles from endosomes. Nature 515: 228-233. doi: 10.1038/nature13846
|
| [11] |
Shimizu H, Kawamura S, Ozaki K (2003) An essential role of Rab5 in uniformity of synaptic vesicle size. J Cell Sci 116: 3583-3590. doi: 10.1242/jcs.00676
|
| [12] |
Satoh AK, O'Tousa JE, Ozaki K, et al. (2005) Rab11 mediates post-Golgi trafficking of rhodopsin to the photosensitive apical membrane of Drosophila photoreceptors. Development 132: 1487-1497. doi: 10.1242/dev.01704
|
| [13] |
Stenmark H (2009) Rab GTPases as coordinators of vesicle traffic. Nat Rev Mol Cell Biol 10: 513-525. doi: 10.1038/nrm2728
|
| [14] |
Polymeropoulos MH, Lavedan C, Leroy E, et al. (1997) Mutation in the alpha-synuclein gene identified in families with Parkinson's disease. Science 276: 2045-2047. doi: 10.1126/science.276.5321.2045
|
| [15] |
Kruger R, Kuhn W, Muller T, et al. (1998) Ala30Pro mutation in the gene encoding alpha-synuclein in Parkinson's disease. Nat Genet 18: 106-108. doi: 10.1038/ng0298-106
|
| [16] |
Jakes R, Spillantini MG, Goedert M (1994) Identification of two distinct synucleins from human brain. FEBS Lett 345: 27-32. doi: 10.1016/0014-5793(94)00395-5
|
| [17] |
Iwai A, Masliah E, Yoshimoto M, et al. (1995) The precursor protein of non-Aβ component of Alzheimer's disease amyloid is a presynaptic protein of the central nervous system. Neuron 14: 467-475. doi: 10.1016/0896-6273(95)90302-X
|
| [18] |
Singleton AB, Farrer M, Johnson J, et al. (2003) alpha-Synuclein locus triplication causes Parkinson's disease. Science 302: 841. doi: 10.1126/science.1090278
|
| [19] |
Jao CC, Der-Sarkissian A, Chen J, et al. (2004) Structure of membrane-bound alpha-synuclein studied by site-directed spin labeling. Proc Natl Acad Sci U S A 101: 8331-8336. doi: 10.1073/pnas.0400553101
|
| [20] |
Abd-Elhadi S, Honig A, Simhi-Haham D, et al. (2015) Total and Proteinase K-Resistant alpha-Synuclein Levels in Erythrocytes, Determined by their Ability to Bind Phospholipids, Associate with Parkinson's Disease. Sci Rep 5: 11120. doi: 10.1038/srep11120
|
| [21] |
Cooper AA, Gitler AD, Cashikar A, et al. (2006) Alpha-synuclein blocks ER-Golgi traffic and Rab1 rescues neuron loss in Parkinson's models. Science 313: 324-328. doi: 10.1126/science.1129462
|
| [22] |
Feany MB, Bender WW (2000) A Drosophila model of Parkinson's disease. Nature 404: 394-398. doi: 10.1038/35006074
|
| [23] |
Periquet M, Fulga T, Myllykangas L, et al. (2007) Aggregated alpha-synuclein mediates dopaminergic neurotoxicity in vivo. J Neurosci 27: 3338-3346. doi: 10.1523/JNEUROSCI.0285-07.2007
|
| [24] |
Chu Y, Morfini GA, Langhamer LB, et al. (2012) Alterations in axonal transport motor proteins in sporadic and experimental Parkinson's disease. Brain 135: 2058-2073. doi: 10.1093/brain/aws133
|
| [25] |
Bayer TA, Jakala P, Hartmann T, et al. (1999) Neural expression profile of alpha-synuclein in developing human cortex. Neuroreport 10: 2799-2803. doi: 10.1097/00001756-199909090-00019
|
| [26] |
Abeliovich A, Schmitz Y, Farinas I, et al. (2000) Mice lacking alpha-synuclein display functional deficits in the nigrostriatal dopamine system. Neuron 25: 239-252. doi: 10.1016/S0896-6273(00)80886-7
|
| [27] | Cabin DE, Shimazu K, Murphy D, et al. (2002) Synaptic vesicle depletion correlates with attenuated synaptic responses to prolonged repetitive stimulation in mice lacking alpha-synuclein. J Neurosci 22: 8797-8807. |
| [28] |
Busch DJ, Oliphint PA, Walsh RB, et al. (2014) Acute increase of alpha-synuclein inhibits synaptic vesicle recycling evoked during intense stimulation. Mol Biol Cell 25: 3926-3941. doi: 10.1091/mbc.E14-02-0708
|
| [29] |
Wang L, Das U, Scott DA, et al. (2014) alpha-synuclein multimers cluster synaptic vesicles and attenuate recycling. Curr Biol 24: 2319-2326. doi: 10.1016/j.cub.2014.08.027
|
| [30] |
Breda C, Nugent ML, Estranero JG, et al. (2015) Rab11 modulates alpha-synuclein-mediated defects in synaptic transmission and behaviour. Hum Mol Genet 24: 1077-1091. doi: 10.1093/hmg/ddu521
|
| [31] |
Paisan-Ruiz C, Jain S, Evans EW, et al. (2004) Cloning of the gene containing mutations that cause PARK8-linked Parkinson's disease. Neuron 44: 595-600. doi: 10.1016/j.neuron.2004.10.023
|
| [32] |
Zimprich A, Biskup S, Leitner P, et al. (2004) Mutations in LRRK2 cause autosomal-dominant parkinsonism with pleomorphic pathology. Neuron 44: 601-607. doi: 10.1016/j.neuron.2004.11.005
|
| [33] |
Nuytemans K, Theuns J, Cruts M, et al. (2010) Genetic etiology of Parkinson disease associated with mutations in the SNCA, PARK2, PINK1, PARK7, and LRRK2 genes: a mutation update. Hum Mutat 31: 763-780. doi: 10.1002/humu.21277
|
| [34] |
Simon-Sanchez J, Schulte C, Bras JM, et al. (2009) Genome-wide association study reveals genetic risk underlying Parkinson's disease. Nat Genet 41: 1308-1312. doi: 10.1038/ng.487
|
| [35] |
Satake W, Nakabayashi Y, Mizuta I, et al. (2009) Genome-wide association study identifies common variants at four loci as genetic risk factors for Parkinson's disease. Nat Genet 41: 1303-1307. doi: 10.1038/ng.485
|
| [36] |
Yao C, Johnson WM, Gao Y, et al. (2013) Kinase inhibitors arrest neurodegeneration in cell and C elegans models of LRRK2 toxicity. Hum Mol Genet 22: 328-344. doi: 10.1093/hmg/dds431
|
| [37] | West AB, Moore DJ, Choi C, et al. (2007) Parkinson's disease-associated mutations in LRRK2 link enhanced GTP-binding and kinase activities to neuronal toxicity. Hum Mol Genet 16: 223-232. |
| [38] |
Smith WW, Pei Z, Jiang H, et al. (2005) Leucine-rich repeat kinase 2 (LRRK2) interacts with parkin, and mutant LRRK2 induces neuronal degeneration. Proc Natl Acad Sci U S A 102: 18676-18681. doi: 10.1073/pnas.0508052102
|
| [39] |
Tong Y, Pisani A, Martella G, et al. (2009) R1441C mutation in LRRK2 impairs dopaminergic neurotransmission in mice. Proc Natl Acad Sci U S A 106: 14622-14627. doi: 10.1073/pnas.0906334106
|
| [40] |
Imai Y, Gehrke S, Wang HQ, et al. (2008) Phosphorylation of 4E-BP by LRRK2 affects the maintenance of dopaminergic neurons in Drosophila. EMBO J 27: 2432-2443. doi: 10.1038/emboj.2008.163
|
| [41] |
Imai Y, Kobayashi Y, Inoshita T, et al. (2015) The Parkinson's Disease-Associated Protein Kinase LRRK2 Modulates Notch Signaling through the Endosomal Pathway. PLoS Genet 11: e1005503. doi: 10.1371/journal.pgen.1005503
|
| [42] |
Rivero-Rios P, Gomez-Suaga P, Fernandez B, et al. (2015) Alterations in late endocytic trafficking related to the pathobiology of LRRK2-linked Parkinson's disease. Biochem Soc Trans 43: 390-395. doi: 10.1042/BST20140301
|
| [43] |
Lee S, Liu HP, Lin WY, et al. (2010) LRRK2 Kinase Regulates Synaptic Morphology through Distinct Substrates at the Presynaptic and Postsynaptic Compartments of the Drosophila Neuromuscular Junction. J Neurosci 30: 16959-16969. doi: 10.1523/JNEUROSCI.1807-10.2010
|
| [44] |
Matta S, Van Kolen K, da Cunha R, et al. (2012) LRRK2 controls an EndoA phosphorylation cycle in synaptic endocytosis. Neuron 75: 1008-1021. doi: 10.1016/j.neuron.2012.08.022
|
| [45] |
Arranz AM, Delbroek L, Van Kolen K, et al. (2015) LRRK2 functions in synaptic vesicle endocytosis through a kinase-dependent mechanism. J Cell Sci 128: 541-552. doi: 10.1242/jcs.158196
|
| [46] |
Piccoli G, Condliffe SB, Bauer M, et al. (2011) LRRK2 controls synaptic vesicle storage and mobilization within the recycling pool. J Neurosci 31: 2225-2237. doi: 10.1523/JNEUROSCI.3730-10.2011
|
| [47] |
Yun HJ, Park J, Ho DH, et al. (2013) LRRK2 phosphorylates Snapin and inhibits interaction of Snapin with SNAP-25. Exp Mol Med 45: e36. doi: 10.1038/emm.2013.68
|
| [48] |
Shin N, Jeong H, Kwon J, et al. (2008) LRRK2 regulates synaptic vesicle endocytosis. Exp Cell Res 314: 2055-2065. doi: 10.1016/j.yexcr.2008.02.015
|
| [49] |
Yun HJ, Kim H, Ga I, et al. (2015) An early endosome regulator, Rab5b, is an LRRK2 kinase substrate. J Biochem 157: 485-495. doi: 10.1093/jb/mvv005
|
| [50] |
Kessels MM, Qualmann B (2002) Syndapins integrate N-WASP in receptor-mediated endocytosis. EMBO J 21: 6083-6094. doi: 10.1093/emboj/cdf604
|
| [51] |
Kim Y, Kim S, Lee S, et al. (2005) Interaction of SPIN90 with dynamin I and its participation in synaptic vesicle endocytosis. J Neurosci 25: 9515-9523. doi: 10.1523/JNEUROSCI.1643-05.2005
|
| [52] |
Soulet F, Yarar D, Leonard M, et al. (2005) SNX9 regulates dynamin assembly and is required for efficient clathrin-mediated endocytosis. Mol Biol Cell 16: 2058-2067. doi: 10.1091/mbc.E04-11-1016
|
| [53] |
Dodson MW, Zhang T, Jiang C, et al. (2012) Roles of the Drosophila LRRK2 homolog in Rab7-dependent lysosomal positioning. Hum Mol Genet 21: 1350-1363. doi: 10.1093/hmg/ddr573
|
| [54] |
Dodson MW, Leung LK, Lone M, et al. (2014) Novel ethyl methanesulfonate (EMS)-induced null alleles of the Drosophila homolog of LRRK2 reveal a crucial role in endolysosomal functions and autophagy in vivo. Dis Model Mech 7: 1351-1363. doi: 10.1242/dmm.017020
|
| [55] |
Esteves AR, M GF, Santos D, et al. (2015) The Upshot of LRRK2 Inhibition to Parkinson's Disease Paradigm. Mol Neurobiol 52: 1804-1820. doi: 10.1007/s12035-014-8980-6
|
| [56] |
Gomez-Suaga P, Rivero-Rios P, Fdez E, et al. (2014) LRRK2 delays degradative receptor trafficking by impeding late endosomal budding through decreasing Rab7 activity. Hum Mol Genet 23: 6779-6796. doi: 10.1093/hmg/ddu395
|
| [57] |
Tong Y, Yamaguchi H, Giaime E, et al. (2010) Loss of leucine-rich repeat kinase 2 causes impairment of protein degradation pathways, accumulation of alpha-synuclein, and apoptotic cell death in aged mice. Proc Natl Acad Sci U S A 107: 9879-9884. doi: 10.1073/pnas.1004676107
|
| [58] |
Alegre-Abarrategui J, Christian H, Lufino MM, et al. (2009) LRRK2 regulates autophagic activity and localizes to specific membrane microdomains in a novel human genomic reporter cellular model. Hum Mol Genet 18: 4022-4034. doi: 10.1093/hmg/ddp346
|
| [59] |
Tong Y, Giaime E, Yamaguchi H, et al. (2012) Loss of leucine-rich repeat kinase 2 causes age-dependent bi-phasic alterations of the autophagy pathway. Mol Neurodegener 7: 2. doi: 10.1186/1750-1326-7-2
|
| [60] |
Vilarino-Guell C, Wider C, Ross OA, et al. (2011) VPS35 mutations in Parkinson disease. Am J Hum Genet 89: 162-167. doi: 10.1016/j.ajhg.2011.06.001
|
| [61] |
Zimprich A, Benet-Pages A, Struhal W, et al. (2011) A mutation in VPS35, encoding a subunit of the retromer complex, causes late-onset Parkinson disease. Am J Hum Genet 89: 168-175. doi: 10.1016/j.ajhg.2011.06.008
|
| [62] |
Nothwehr SF, Bruinsma P, Strawn LA (1999) Distinct domains within Vps35p mediate the retrieval of two different cargo proteins from the yeast prevacuolar/endosomal compartment. Mol Biol Cell 10: 875-890. doi: 10.1091/mbc.10.4.875
|
| [63] |
Korolchuk VI, Schutz MM, Gomez-Llorente C, et al. (2007) Drosophila Vps35 function is necessary for normal endocytic trafficking and actin cytoskeleton organisation. J Cell Sci 120: 4367-4376. doi: 10.1242/jcs.012336
|
| [64] |
Kumar KR, Weissbach A, Heldmann M, et al. (2012) Frequency of the D620N Mutation in VPS35 in Parkinson Disease. Arch Neurol 69: 1360-1364. doi: 10.1001/archneurol.2011.3367
|
| [65] |
Ando M, Funayama M, Li Y, et al. (2012) VPS35 mutation in Japanese patients with typical Parkinson's disease. Mov Disord 27: 1413-1417. doi: 10.1002/mds.25145
|
| [66] |
Follett J, Norwood SJ, Hamilton NA, et al. (2014) The Vps35 D620N mutation linked to Parkinson's disease disrupts the cargo sorting function of retromer. Traffic 15: 230-244. doi: 10.1111/tra.12136
|
| [67] |
Zavodszky E, Seaman MN, Moreau K, et al. (2014) Mutation in VPS35 associated with Parkinson's disease impairs WASH complex association and inhibits autophagy. Nat Commun 5: 3828-. doi: 10.1038/ncomms4828
|
| [68] |
McGough IJ, Steinberg F, Jia D, et al. (2014) Retromer binding to FAM21 and the WASH complex is perturbed by the Parkinson disease-linked VPS35(D620N) mutation. Curr Biol 24: 1670-1676. doi: 10.1016/j.cub.2014.06.024
|
| [69] |
Temkin P, Lauffer B, Jager S, et al. (2011) SNX27 mediates retromer tubule entry and endosome-to-plasma membrane trafficking of signalling receptors. Nat Cell Biol 13: 715-721. doi: 10.1038/ncb2252
|
| [70] |
Zech T, Calaminus SD, Caswell P, et al. (2011) The Arp2/3 activator WASH regulates alpha5beta1-integrin-mediated invasive migration. J Cell Sci 124: 3753-3759. doi: 10.1242/jcs.080986
|
| [71] |
Vilarino-Guell C, Rajput A, Milnerwood AJ, et al. (2014) DNAJC13 mutations in Parkinson disease. Hum Mol Genet 23: 1794-1801. doi: 10.1093/hmg/ddt570
|
| [72] |
Popoff V, Mardones GA, Bai SK, et al. (2009) Analysis of Articulation Between Clathrin and Retromer in Retrograde Sorting on Early Endosomes. Traffic 10: 1868-1880. doi: 10.1111/j.1600-0854.2009.00993.x
|
| [73] |
Freeman CL, Hesketh G, Seaman MNJ (2014) RME-8 coordinates the activity of the WASH complex with the function of the retromer SNX dimer to control endosomal tubulation. J Cell Sci 127: 2053-2070. doi: 10.1242/jcs.144659
|
| [74] |
Munsie LN, Milnerwood AJ, Seibler P, et al. (2015) Retromer-dependent neurotransmitter receptor trafficking to synapses is altered by the Parkinson's disease VPS35 mutation p.D620N. Hum Mol Genet 24: 1691-1703. doi: 10.1093/hmg/ddu582
|
| [75] |
Wang HS, Toh J, Ho P, et al. (2014) In vivo evidence of pathogenicity of VPS35 mutations in the Drosophila. Mol Brain 7: 73. doi: 10.1186/s13041-014-0073-y
|
| [76] |
Wen L, Tang FL, Hong Y, et al. (2011) VPS35 haploinsufficiency increases Alzheimer's disease neuropathology. J Cell Biol 195: 765-779. doi: 10.1083/jcb.201105109
|
| [77] |
MacLeod DA, Rhinn H, Kuwahara T, et al. (2013) RAB7L1 interacts with LRRK2 to modify intraneuronal protein sorting and Parkinson's disease risk. Neuron 77: 425-439. doi: 10.1016/j.neuron.2012.11.033
|
| [78] |
Beilina A, Rudenko IN, Kaganovich A, et al. (2014) Unbiased screen for interactors of leucine-rich repeat kinase 2 supports a common pathway for sporadic and familial Parkinson disease. Proc Natl Acad Sci U S A 111: 2626-2631. doi: 10.1073/pnas.1318306111
|
| [79] |
Linhart R, Wong SA, Cao J, et al. (2014) Vacuolar protein sorting 35 (Vps35) rescues locomotor deficits and shortened lifespan in Drosophila expressing a Parkinson's disease mutant of Leucine-Rich Repeat Kinase 2 (LRRK2). Mol Neurodegener 9: 23. doi: 10.1186/1750-1326-9-23
|
| [80] |
Edvardson S, Cinnamon Y, Ta-Shma A, et al. (2012) A deleterious mutation in DNAJC6 encoding the neuronal-specific clathrin-uncoating co-chaperone auxilin, is associated with juvenile parkinsonism. PLoS One 7: e36458. doi: 10.1371/journal.pone.0036458
|
| [81] |
Koroglu C, Baysal L, Cetinkaya M, et al. (2013) DNAJC6 is responsible for juvenile parkinsonism with phenotypic variability. Parkinsonism Relat Disord 19: 320-324. doi: 10.1016/j.parkreldis.2012.11.006
|
| [82] |
Fotin A, Cheng YF, Sliz P, et al. (2004) Molecular model for a complete clathrin lattice from electron cryomicroscopy. Nature 432: 573-579. doi: 10.1038/nature03079
|
| [83] |
Young JC, Barral JM, Hartl FU (2003) More than folding: localized functions of cytosolic chaperones. Trends Biochem Sci 28: 541-547. doi: 10.1016/j.tibs.2003.08.009
|
| [84] |
Pankratz N, Wilk JB, Latourelle JC, et al. (2009) Genomewide association study for susceptibility genes contributing to familial Parkinson disease. Hum Genet 124: 593-605. doi: 10.1007/s00439-008-0582-9
|
| [85] |
Yim YI, Sun T, Wu LG, et al. (2010) Endocytosis and clathrin-uncoating defects at synapses of auxilin knockout mice. Proc Natl Acad Sci U S A 107: 4412-4417. doi: 10.1073/pnas.1000738107
|
| [86] |
Dumitriu A, Pacheco CD, Wilk JB, et al. (2011) Cyclin-G-associated kinase modifies alpha-synuclein expression levels and toxicity in Parkinson's disease: results from the GenePD Study. Hum Mol Genet 20: 1478-1487. doi: 10.1093/hmg/ddr026
|
| [87] |
Krebs CE, Karkheiran S, Powell JC, et al. (2013) The Sac1 domain of SYNJ1 identified mutated in a family with early-onset progressive Parkinsonism with generalized seizures. Hum Mutat 34: 1200-1207. doi: 10.1002/humu.22372
|
| [88] |
Nalls MA, Pankratz N, Lill CM, et al. (2014) Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson's disease. Nat Genet 46: 989-993. doi: 10.1038/ng.3043
|
| [89] | Luo WJ, Chang A (1997) Novel genes involved in endosomal traffic in yeast revealed by suppression of a targeting-defective plasma membrane ATPase mutant. Mol Biol Cell 8: 1779-1779. |
| [90] |
Verstreken P, Koh TW, Schulze KL, et al. (2003) Synaptojanin is recruited by Endophilin to promote synaptic vesicle uncoating. Neuron 40: 733-748. doi: 10.1016/S0896-6273(03)00644-5
|
| [91] |
Harris TW, Hartwieg E, Horvitz HR, et al. (2000) Mutations in synaptojanin disrupt synaptic vesicle recycling. J Cell Biol 150: 589-599. doi: 10.1083/jcb.150.3.589
|
| [92] |
Schuske KR, Richmond JE, Matthies DS, et al. (2003) Endophilin is required for synaptic vesicle endocytosis by localizing synaptojanin. Neuron 40: 749-762. doi: 10.1016/S0896-6273(03)00667-6
|
| [93] |
Periquet M, Corti O, Jacquier S, et al. (2005) Proteomic analysis of parkin knockout mice: alterations in energy metabolism, protein handling and synaptic function. J Neurochem 95: 1259-1276. doi: 10.1111/j.1471-4159.2005.03442.x
|
| [94] |
Kitada T, Pisani A, Porter DR, et al. (2007) Impaired dopamine release and synaptic plasticity in the striatum of PINK1-deficient mice. Proc Natl Acad Sci U S A 104: 11441-11446. doi: 10.1073/pnas.0702717104
|
| [95] |
Morais VA, Verstreken P, Roethig A, et al. (2009) Parkinson's disease mutations in PINK1 result in decreased Complex I activity and deficient synaptic function. EMBO Mol Med 1: 99-111. doi: 10.1002/emmm.200900006
|
| [96] |
Vincent A, Briggs L, Chatwin GF, et al. (2012) parkin-induced defects in neurophysiology and locomotion are generated by metabolic dysfunction and not oxidative stress. Hum Mol Genet 21: 1760-1769. doi: 10.1093/hmg/ddr609
|
| [97] |
Shiba-Fukushima K, Inoshita T, Hattori N, et al. (2014) PINK1-Mediated Phosphorylation of Parkin Boosts Parkin Activity in Drosophila. PLoS Genet 10: e1004391. doi: 10.1371/journal.pgen.1004391
|
| [98] |
Braschi E, Goyon V, Zunino R, et al. (2010) Vps35 Mediates Vesicle Transport between the Mitochondria and Peroxisomes. Curr Biol 20: 1310-1315. doi: 10.1016/j.cub.2010.05.066
|
| [99] |
Tang FL, Liu W, Hu JX, et al. (2015) VPS35 Deficiency or Mutation Causes Dopaminergic Neuronal Loss by Impairing Mitochondrial Fusion and Function. Cell Rep 12: 1631-1643. doi: 10.1016/j.celrep.2015.08.001
|
| [100] |
Cali T, Ottolini D, Negro A, et al. (2012) alpha-Synuclein Controls Mitochondrial Calcium Homeostasis by Enhancing Endoplasmic Reticulum-Mitochondria Interactions. J Biol Chem 287: 17914-17929. doi: 10.1074/jbc.M111.302794
|
| [101] |
Nakamura K, Nemani VM, Azarbal F, et al. (2011) Direct Membrane Association Drives Mitochondrial Fission by the Parkinson Disease-associated Protein alpha-Synuclein. J Biol Chem 286: 20710-20726. doi: 10.1074/jbc.M110.213538
|
| [102] |
Wong YC, Holzbaur EL (2014) Optineurin is an autophagy receptor for damaged mitochondria in parkin-mediated mitophagy that is disrupted by an ALS-linked mutation. Proc Natl Acad Sci U S A 42: E4439-48. doi: 10.1073/pnas.1405752111
|
| [103] |
Guardia-Laguarta C, Area-Gomez E, Rub C, et al. (2014) alpha-Synuclein Is Localized to Mitochondria-Associated ER Membranes. J Neurosci 34: 249-259. doi: 10.1523/JNEUROSCI.2507-13.2014
|
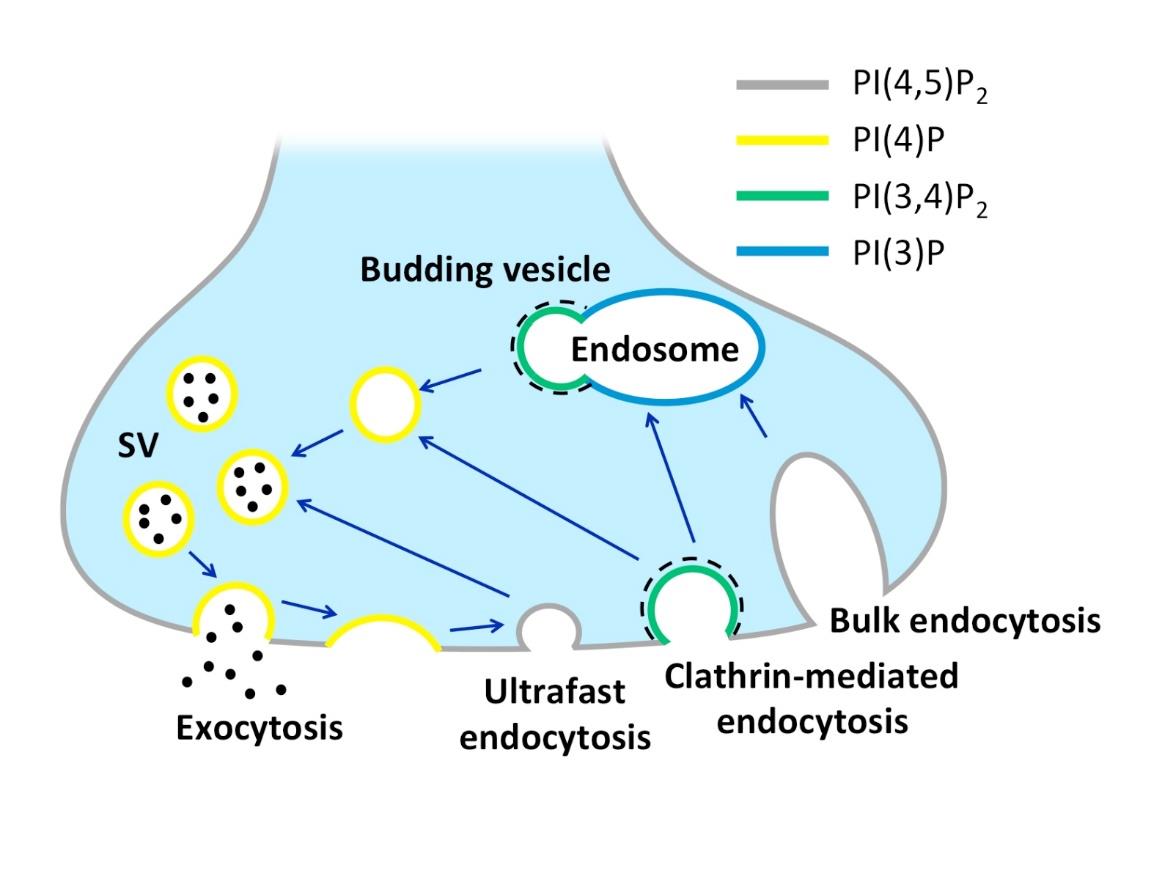
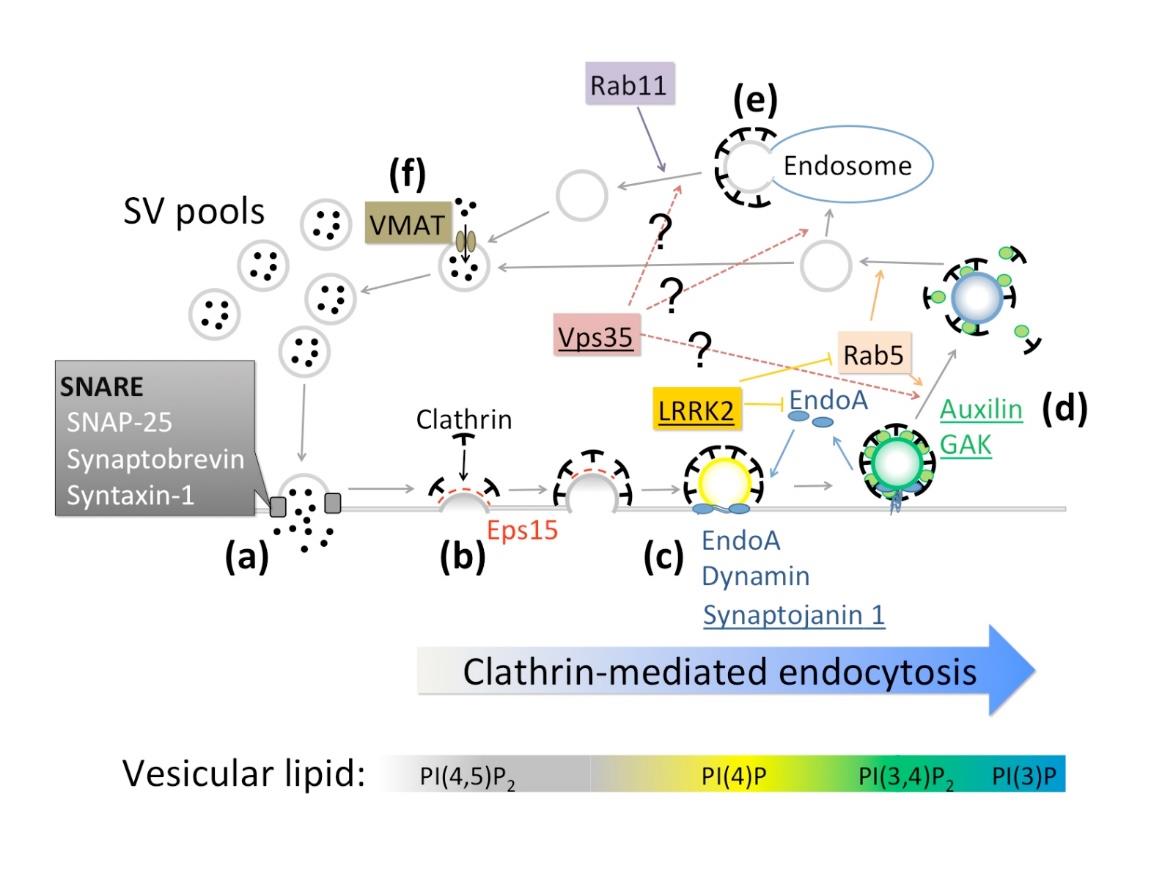
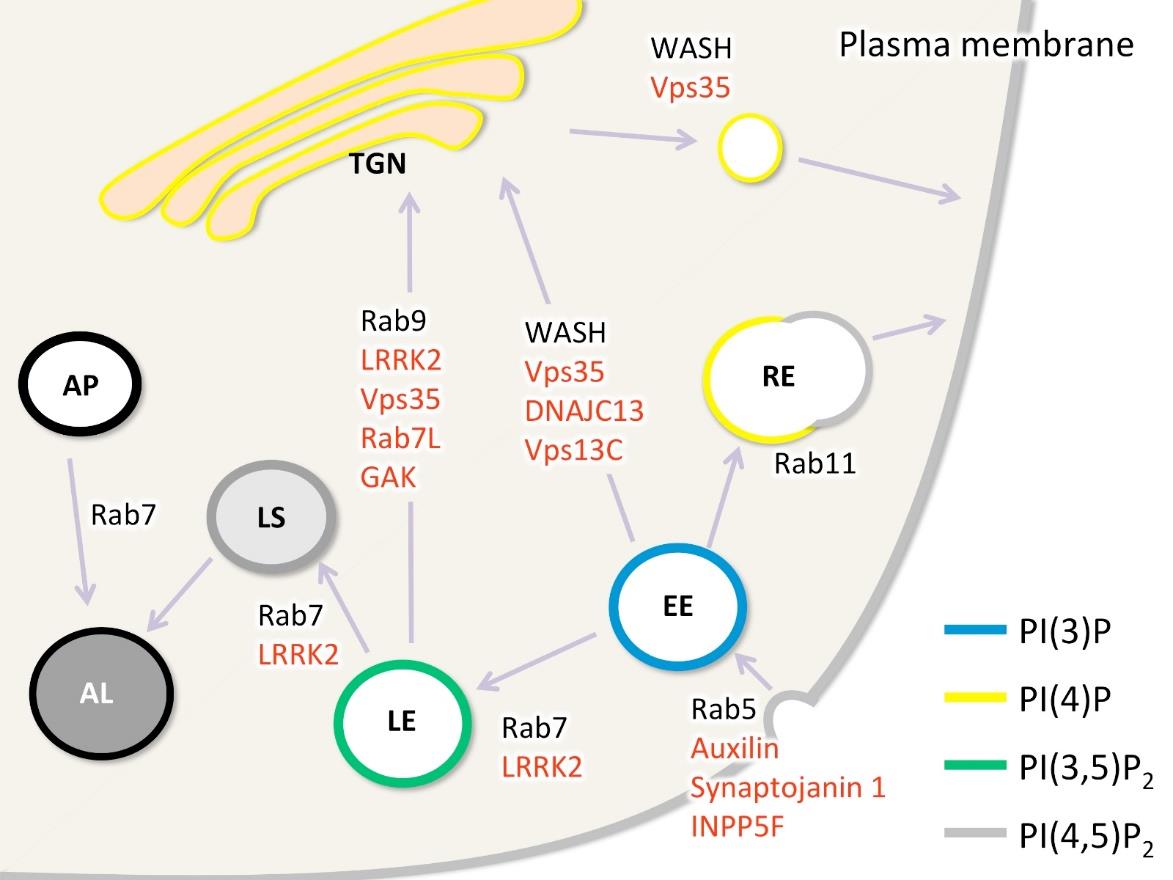
Figures(3) / Tables(1)

Tsuyoshi Inoshita, Yuzuru Imai. Regulation of vesicular trafficking by Parkinson's disease-associated genes[J]. AIMS Molecular Science, 2015, 2(4): 461-475. doi: 10.3934/molsci.2015.4.461







 DownLoad:
DownLoad: