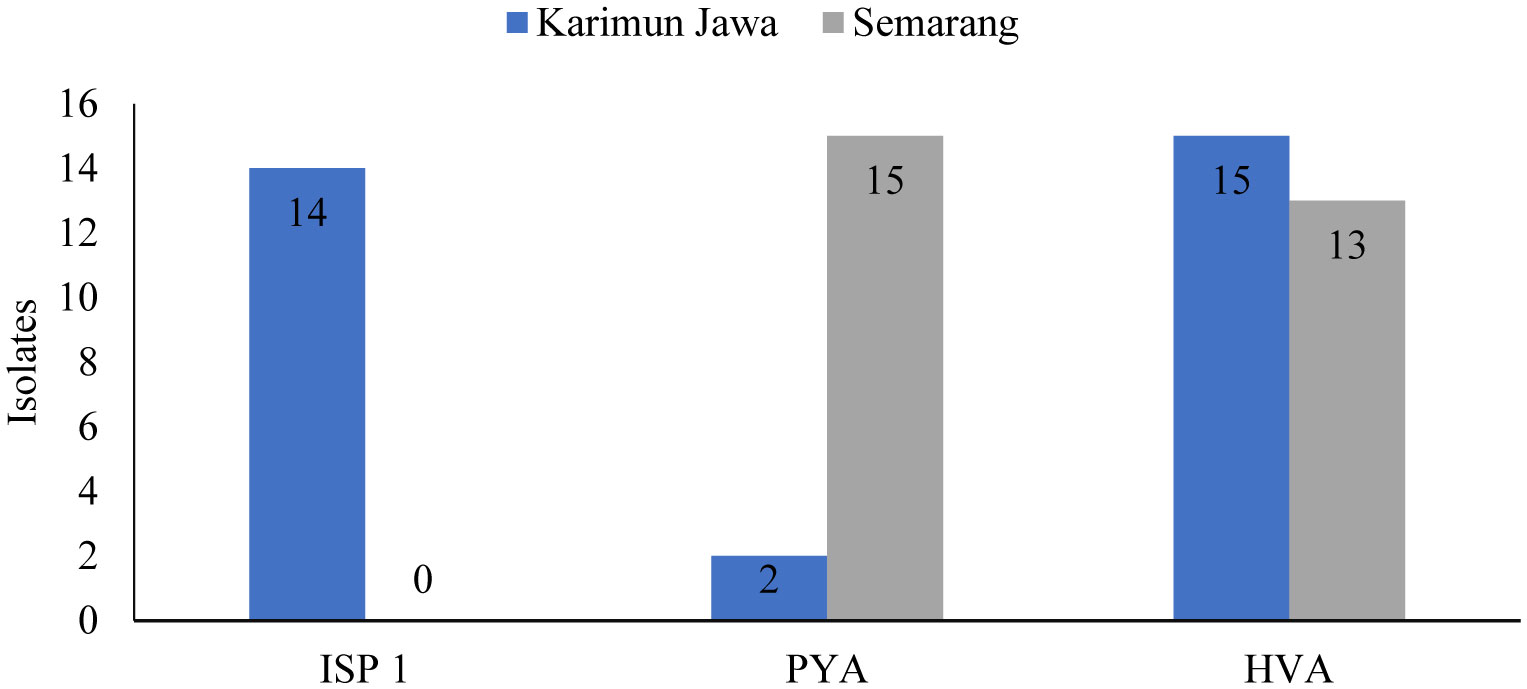
Infection caused by pathogenic fungal species is one of the most challenging disease to be tackled today. The antifungal bacteria candidate can be found in terrestrial as well as aquatic ecosystems, with mangrove forests being one of them. The purpose of this study is to obtain candidate isolates of antifungal strains with a detection approach and gene mapping simulation of bioactive compounds producers and screening to determine qualitative antifungal activity. The research will be carried out by collecting sediment samples from the mangrove ecosystems of Karimunjawa and Mangkang sub-district of Semarang city, isolating and purifying bacteria with Humic Acid Vitamin Agar (HVA), International Streptomyces Project 1 (ISP 1) and Zobell (Marine Agar). added with antibiotics, qualitative antifungal ability screening of each isolate obtained, detection of the presence of PKS gene and NRPS using special primers using the Polymerase Chain Reaction (PCR) method, and molecular identification of each isolate by 16s rRNA sequencing method. Of the total 59 isolates produced from the sample isolation process, 31 isolates from Karimunjawa sediments and 8 isolates from Semarang sediments showed activity against test pathogenic bacteria, namely Candida albicans, Trichoderma sp., and Aspergillus niger. Detection of Biosynthethic Gene Cluster (BGC) showed that the genes encoding secondary metabolites (NRPS, PKS 1 and PKS 2) were detected in KI 2-2 isolates from Karimunjawa. NRPS were detected only in isolates SP 3-9, SH 3-4, KI 1-6, KI 2-2, KI 2-4. The secondary metabolite-encoding gene, PKS1, was detected in isolates SP 3-5, SP 3-8, KI 2-2. PKS II genes were detected only on isolates SP 2-4, SH 3-8, KI 1-6, KI 2-2, and KI 2-4. Isolate SP 3-5 was revealed as Pseudomonas aeruginosa (93.14%), isolate SP 2-4 was Zhouia amylolytica strain HN-181 (100%) and isolate SP 3-8 was P. aeruginosa strain QK -2 (100%).
Citation: Delianis Pringgenies, Wilis Ari Setyati. Antifungal strains and gene mapping of secondary metabolites in mangrove sediments from Semarang city and Karimunjawa islands, Indonesia[J]. AIMS Microbiology, 2021, 7(4): 499-512. doi: 10.3934/microbiol.2021030
Infection caused by pathogenic fungal species is one of the most challenging disease to be tackled today. The antifungal bacteria candidate can be found in terrestrial as well as aquatic ecosystems, with mangrove forests being one of them. The purpose of this study is to obtain candidate isolates of antifungal strains with a detection approach and gene mapping simulation of bioactive compounds producers and screening to determine qualitative antifungal activity. The research will be carried out by collecting sediment samples from the mangrove ecosystems of Karimunjawa and Mangkang sub-district of Semarang city, isolating and purifying bacteria with Humic Acid Vitamin Agar (HVA), International Streptomyces Project 1 (ISP 1) and Zobell (Marine Agar). added with antibiotics, qualitative antifungal ability screening of each isolate obtained, detection of the presence of PKS gene and NRPS using special primers using the Polymerase Chain Reaction (PCR) method, and molecular identification of each isolate by 16s rRNA sequencing method. Of the total 59 isolates produced from the sample isolation process, 31 isolates from Karimunjawa sediments and 8 isolates from Semarang sediments showed activity against test pathogenic bacteria, namely Candida albicans, Trichoderma sp., and Aspergillus niger. Detection of Biosynthethic Gene Cluster (BGC) showed that the genes encoding secondary metabolites (NRPS, PKS 1 and PKS 2) were detected in KI 2-2 isolates from Karimunjawa. NRPS were detected only in isolates SP 3-9, SH 3-4, KI 1-6, KI 2-2, KI 2-4. The secondary metabolite-encoding gene, PKS1, was detected in isolates SP 3-5, SP 3-8, KI 2-2. PKS II genes were detected only on isolates SP 2-4, SH 3-8, KI 1-6, KI 2-2, and KI 2-4. Isolate SP 3-5 was revealed as Pseudomonas aeruginosa (93.14%), isolate SP 2-4 was Zhouia amylolytica strain HN-181 (100%) and isolate SP 3-8 was P. aeruginosa strain QK -2 (100%).

| [1] |
Ariyanto D, Bengen DG, Prartono T, et al. (2020) Distribution and abundance of Cerithideopsilla djadjariensis (Martin 1899) (potamididae) on avicennia marina in Rembang, Central Java, Indonesia. Egypt J Aquat Biol Fish 24: 323-332. doi: 10.21608/ejabf.2020.95329

|
| [2] | Ariyanto D, Gunawan H, Puspitasari D, et al. (2019) The differences of the elements content in Rhizophora mucronata leaves from Asahan Regency, North Sumatra, Indonesia. Polish J Nat Sci 34: 481-491. |
| [3] |
Ningsih SS, Ariyanto D, Puspitasari D, et al. (2020) The amino acid contents in mangrove Rhizophora mucronata leaves in Asahan, North Sumatra, Indonesia. E3S Web Conf 151: 1-3. doi: 10.1051/e3sconf/202015101047
|
| [4] |
Pringgenies D, Setyati WA, Djunaedi A, et al. (2021) Exploration of antimicrobial potency of mangrove symbiont against multi-drug resistant bacteria. J Ilm Perikan dan Kelaut 12: 222-232. doi: 10.20473/jipk.v13i2.26199
|
| [5] |
Ariyanto D, Bengen DG, Prartono T, et al. (2018) Short Communication: The relationship between content of particular metabolites of fallen mangrove leaves and the rate at which the leaves decompose over time. Biodiversitas 19: 700-705. doi: 10.13057/biodiv/d190304
|
| [6] |
Shen N, Liang Z, Liu Q, et al. (2020) Antifungal secondary metabolites isolated from mangrove rhizosphere soil-derived penicillium fungi. J Ocean Univ China 19: 717-721. doi: 10.1007/s11802-020-4360-1
|
| [7] | Da X, Nishiyama Y, Tie D, et al. (2019) Antifungal activity and mechanism of action of Ou-gon (Scutellaria root extract) components against pathogenic fungi. Sci Rep 9: 1-12. |
| [8] |
Azman AS, Othman I, Velu SS, et al. (2015) Mangrove rare actinobacteria: Taxonomy, natural compound, and discovery of bioactivity. Front Microbiol 6: 1-15. doi: 10.3389/fmicb.2015.00856
|
| [9] |
Sharma P, Thakur D (2020) Antimicrobial biosynthetic potential and diversity of culturable soil actinobacteria from forest ecosystems of Northeast India. Sci Rep 10: 1-18. doi: 10.1038/s41598-019-56847-4
|
| [10] |
Sekurova ON, Schneider O, Zotchev SB (2019) Novel bioactive natural products from bacteria via bioprospecting, genome mining and metabolic engineering. Microb Biotechnol 12: 828-44. doi: 10.1111/1751-7915.13398
|
| [11] |
Davies-Bolorunduro OF, Adeleye IA, Akinleye MO, et al. (2019) Wang PG. Anticancer potential of metabolic compounds from marine actinomycetes isolated from Lagos Lagoon sediment. J Pharm Anal 9: 201-208. doi: 10.1016/j.jpha.2019.03.004
|
| [12] | Li Q, Chen X, Jiang Y, et al. (2016) Morphological Identification of Actinobacteria, In: Dharumadurai Dhanasekaran, Yi Jiang. Actinobacteria-Basics and Biotechnological Applications New York: 60-84. |
| [13] |
Hayakawa M, Nonomura H (1987) Humic acid-vitamin agar, a new medium for the selective isolation of soil actinomycetes. J Ferment Technol 65: 501-509. doi: 10.1016/0385-6380(87)90108-7
|
| [14] |
Messaoudi O, Wink J (2020) Diversity of actinobacteria isolated from date palms rhizosphere and saline environments : isolation, identification and biological activity evaluation. Microorganisms 8: 1853. doi: 10.3390/microorganisms8121853
|
| [15] |
Radjasa OK, Martens T, Grossart HP, et al. (2005) Antibacterial property of a coral-associated bacterium Pseudoalteromonas luteoviolacea against shrimp pathogenic Vibrio harveyi (In Vitro Study). HAYATI J Biosci 12: 77-81. doi: 10.1016/S1978-3019(16)30329-1
|
| [16] |
El Samak M, Solyman SM, Hanora A (2018) Antimicrobial activity of bacteria isolated from Red Sea marine invertebrates. Biotechnol Reports 19: e00275. doi: 10.1016/j.btre.2018.e00275
|
| [17] | Prescott LM, Harley JP, Klein D (1999) Microbiology New York: The McGraw-Hill Companies, 685. |
| [18] | Majewski M, Kozlowska A, Thoene M, et al. (2016) Overview of the role of vitamins and minerals on the kynurenine. J Physiol Pharmacol 67: 3-19. |
| [19] |
Anggelina AC, Pringgenies D, Setyati WA (2021) Presence of biosynthetic gene clusters (NRPS/PKS) in actinomycetes of mangrove sediment in Semarang and Karimunjawa, Indonesia. Environ Nat Resour J 19: 391-401. doi: 10.32526/ennrj/19/202100050
|
| [20] |
Thatoi H, Behera BC, Mishra RR, et al. (2013) Biodiversity and biotechnological potential of microorganisms from mangrove ecosystems: A review. Ann Microbiol 63: 1-19. doi: 10.1007/s13213-012-0442-7
|
| [21] | Pringgenies D, Widiyadmi R, Ariyanto D, et al. (2019) Organic compost production from bacterial consortium of mangrove leaf litter. J Pengelolaan Perair 2: 29-26. |
| [22] |
Moradali MF, Ghods S, Rehm BHA (2017) Pseudomonas aeruginosa lifestyle: A paradigm for adaptation, survival, and persistence. Front Cell Infect Microbiol 7: 1-29. doi: 10.3389/fcimb.2017.00039
|
| [23] | Murcia NR, Waridel P, Maspoli A, et al. (2015) The Pseudomonas aeruginosa antimetabolite L-2-amino-4-methoxy-trans-3-butenoic acid (AMB) is made from glutamate and two alanine residues via a thiotemplate-linked tripeptide precursor. Front Microbiol 6: 1-13. |
| [24] | Pringgenies D, Diponegoro U, Ridlo A, et al. (2011) Production of phenazine pigments from marine symbiotic bacteria in gastropod Cerithidea sp. with different Growth Media. Conf Int Converence Nat Sci 2011 153-162. |
| [25] |
Ho Sui SJ, Lo R, Fernandes AR, et al. (2012) Raloxifene attenuates Pseudomonas aeruginosa pyocyanin production and virulence. Int J Antimicrob Agents 40: 246-251. doi: 10.1016/j.ijantimicag.2012.05.009
|
| [26] |
Ciofu O, Tolker-Nielsen T (2019) Tolerance and resistance of pseudomonas aeruginosabiofilms to antimicrobial agents-how P. aeruginosa Can escape antibiotics. Front Microbiol 10: 1-15. doi: 10.3389/fmicb.2019.00913
|
| [27] | Juliadiningtyas AD, Pringgenies D, Heriyanto, et al. (2018) Preliminary investigation of the carotenoid composition of Erythrobacter sp. Strain KJ5 by high-performance liquid chromatography and mass spectrometry. Philipp J Sci 147: 91-98. |
| [28] | Kanesaki Y, Setiyono E, Pringgenies D, et al. (2019) Complete genome sequence of the marine bacterium Erythrobacter flavus Strain KJ5. Microbiol Resosrces Announc 8: 1-2. |
| [29] |
Setiyono E, Heriyanto, Pringgenies D, et al. (2019) Sulfur-containing carotenoids from a marine coral symbiont Erythrobacter flavus Strain KJ5. Mar Drugs 17: 1-15. doi: 10.3390/md17060349
|
| [30] |
Liu ZP, Wang BJ, Dai X, et al. (2006) Zhouia amylolytica gen. nov., sp. nov., a novel member of the family Flavobacteriaceae isolated from sediment of the South China Sea. Int J Syst Evol Microbiol 56: 2825-2829. doi: 10.1099/ijs.0.64587-0
|
| [31] | Yang CW, Chao WL, Hsieh CY, et al. (2019) Biodegradation of sulfamethoxazole in milkfish (Chanos chanos) pond sediments. Appl Sci 9: 1-16. |

Figures(2) / Tables(7)

Delianis Pringgenies, Wilis Ari Setyati. Antifungal strains and gene mapping of secondary metabolites in mangrove sediments from Semarang city and Karimunjawa islands, Indonesia[J]. AIMS Microbiology, 2021, 7(4): 499-512. doi: 10.3934/microbiol.2021030







 DownLoad:
DownLoad: