Citation: George P. Stamou, Sotiris Konstadinou, Nikolaos Monokrousos, Anna Mastrogianni, Michalis Orfanoudakis, Christos Hassiotis, Urania Menkissoglu-Spiroudi, Despoina Vokou, Efimia M. Papatheodorou. The effects of arbuscular mycorrhizal fungi and essential oil on soil microbial community and N-related enzymes during the fungal early colonization phase[J]. AIMS Microbiology, 2017, 3(4): 938-959. doi: 10.3934/microbiol.2017.4.938

| [1] |
Watt M, Kirkegaard JA, Passioura JB (2006) Rhizosphere biology and crop productivity-a review. Soil Res 44: 299–317. doi: 10.1071/SR05142

|
| [2] |
Tahat MM, Sijam K (2012) Arbuscular mycorrhizal fungi and plant root exudates bio-communications in the rhizosphere. Afr J Microbiol Res 6: 7295–7301. doi: 10.5897/AJMR12.2250
|
| [3] |
Miransari M (2011) Interactions between arbuscular mycorrhizal fungi and soil bacteria. Appl Microbiol Biot 89: 917–930. doi: 10.1007/s00253-010-3004-6
|
| [4] | Schüβler A, Walker C (2010) The Glomeromycota: A species list with new families and new genera. Libraries at the Royal Botanic Garden Edinburgh, Kew, Botanische Staatssammlung Munich and Oregon State University. Available from: www.amf-phylogeny.com. |
| [5] |
Velázquez S, Cabello M (2011) Occurrence and diversity of arbuscular mycorrhizal fungi in trap cultures from El Palmer National Park soils. Eur J Soil Biol 47: 230–235. doi: 10.1016/j.ejsobi.2011.05.002
|
| [6] |
Veresoglou SD, Chen B, Rillig MC (2012) Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biol Biochem 46: 53–62. doi: 10.1016/j.soilbio.2011.11.018
|
| [7] |
Lioussanne L, Perreault F, Jolicoeur M, et al. (2010) The bacterial community of tomato rhizosphere is modified by inoculation with arbuscular mycorrhizal fungi but unaffected by soil enrichment with mycorrhizal root exudates or inoculation with Phytophthora nicotianae. Soil Biol Biochem 42: 473–483. doi: 10.1016/j.soilbio.2009.11.034
|
| [8] |
Vazquez MM, Cesar S, Azcon R, et al. (2000) Interactions between arbuscular mycorrhizal fungi and other microbial inoculants (Azospirillum, Pseudomonas, Trichoderma) and their effects on microbial population and enzyme activities in the rhizosphere of maize plants. Appl Soil Ecol 15: 261–272. doi: 10.1016/S0929-1393(00)00075-5
|
| [9] |
Bona E, Lingua G, Manassero P, et al. (2015) AM fungi and PGP pseudomonads increase flowering, fruit production, and vitamin content in strawberry grown at low nitrogen and phosphorus levels. Mycorrhiza 25: 181–193. doi: 10.1007/s00572-014-0599-y
|
| [10] |
Lazcano C, Barrios-Masias FH, Jackson LE (2014) Arbuscular mycorrhizal effects on plant water relations and soil greenhouse gas emissions under changing moisture regimes. Soil Biol Biochem 74: 184–192. doi: 10.1016/j.soilbio.2014.03.010
|
| [11] |
du Jardin P (2015) Plant biostimulants: Definition, concept, main categories and regulation. Sci Hortic 196: 3–14. doi: 10.1016/j.scienta.2015.09.021
|
| [12] | Rouphael Y, Cardarelli M, Colla G (2015)Role of arbuscular mycorrhizal fungi in alleviating the adverse effects of acidity and aluminium toxicity in zucchini squash. Sci Hortic 188: 97–105. |
| [13] |
Rouphael Y, Cardarelli M, di Mattia E, et al. (2010) Enhancement of alkalinity tolerance in two cucumber genotypes inoculated with an arbuscular mycorrhizal biofertilizer containing Glomus intraradices. Biol Fertil Soils 46: 499–509. doi: 10.1007/s00374-010-0457-9
|
| [14] |
Inderjit (1996) Plant phenolics in allelopathy. Bot Rev 62: 186–202. doi: 10.1007/BF02857921
|
| [15] | Vokou D (2007) Allelochemicals, allelopathy and essential oils: A field in search of definitions and structure. Allelopathy J 19: 119–135. |
| [16] |
Papatheodorou EM, Margariti C, Vokou D (2014) Effects of the two carvone enantiomers on soil enzymes involved in C, P and N cycles. J Biol Res-Thessaloniki 21: 7. doi: 10.1186/2241-5793-21-7
|
| [17] |
Kadoglidou K, Lagopodi A, Karamanoli K, et al. (2011) Inhibitory and stimulatory effects of essential oils and individual monoterpenoids on growth and sporulation of four soil-borne fungal isolates of Aspergillus terreus, Fusarium oxysporum, Penicillium expansum, and Verticillium dahlia. Eur J Plant Pathol 130: 297–309. doi: 10.1007/s10658-011-9754-x
|
| [18] | Rasoul MAA, Marei GIK, Abdelgaleil SAM (2012) Evaluation of antibacterial properties and biochemical effects of monoterpenes on plant pathogenic bacteria. Afr J Microbiol Res 6: 3667–3672. |
| [19] |
Vokou D, Chalkos D, Karamanlidou G, et al. (2002) Activation of soil respiration and shift of the microbial population balance in soil as a response to Lavandula stoechas essential oil. J Chem Ecol 28: 755–768. doi: 10.1023/A:1015236709767
|
| [20] |
Hassiotis CN (2010) Evaluation of essential oil antifungal activity against mycorrhizal fungi-the case of Laurus nobilis essential oil. Isr J Ecol Evol 56: 35–54. doi: 10.1560/IJEE.56.1.35
|
| [21] |
Hassiotis CN, Dina EI (2011) The effects of laurel (Laurus nobilis L.) on development of two mycorrhizal fungi. Int Biodeter Biodegr 65: 628–634. doi: 10.1016/j.ibiod.2011.03.006
|
| [22] | Pino O, Sanchez Y, Rojas MM (2013) Plant secondary metabolites as an alternative in pest management. I: background, research approaches and trends. Revista De Proteccion Vegetal 28: 81–94. |
| [23] | Kouassi KH, Bajji M, Zhiri A, et al. (2010) Evaluation of three essential oils as potential sources of botanical fungicides. Commun Agr Appl Biol Sci 75: 525–529. |
| [24] |
Wang MY, Xia RX, Wu QS, et al. (2007) Influence of arbuscular mycorrhizal fungi on microbes and enzymes of soils from different cultivated densities of red clover. Ann Microbiol 57: 1–7. doi: 10.1007/BF03175042
|
| [25] |
Nottingham AT, Turner BL, Winter K, et al. (2013) Root and arbuscular mycorrhizal mycelial interactions with soil microorganisms in lowland tropical forest. FEMS Microbiol Ecol 85: 37–50. doi: 10.1111/1574-6941.12096
|
| [26] | Garcia-Garrido J, Vierheiling H (2009) From a germinating spore to an established arbuscular mycorrhiza, In: Khasa D, Piche Y, Coughlan A, Advances in mycorrhizal science and technologies, Council National Research of Canada. |
| [27] |
Seddas PMA, Arias CM, Arnould C, et al. (2009) Symbiosis-related plant genes modulate molecular responses in an arbuscular mycorrhizal fungus during early root interactions. Mol Plant Microbe In 22: 341–351. doi: 10.1094/MPMI-22-3-0341
|
| [28] |
Larose G, Chênevert R, Moutoglis P, et al. (2002) Flavonoid levels in roots of Medicago sativa are modulated by the developmental stage of the symbiosis and the root colonizing arbuscular mycorrhizal fungus. J Plant Physiol 159: 1329–1339. doi: 10.1078/0176-1617-00896
|
| [29] |
Piotrowski JS, Morford SL, Rillig MC (2008) Inhibition of colonization by a native arbuscular mycorrhizal fungal community via Populus trichocarpa litter, litter extract, and soluble phenolic compounds. Soil Biol Biochem 40: 709–717. doi: 10.1016/j.soilbio.2007.10.005
|
| [30] | Al-Tawaha A, Al-Karaki G, Massadeh A (2013) Comparative response of essential oil composition, antioxidant activity and phenolic contents spearmint (Mentha spicata L.) under protected soilless vs. open field. Adv Environ Biol 7: 902–910. |
| [31] | Bimakr M, Rahman RA, Ganjloo A, et al. (2011) Optimization of supercritical carbon dioxide extraction of bioactive flavonoid compounds from spearmint (Mentha spicata L.) leaves by using response surface methodology. Food Bioprocess Tech 5: 912–920. |
| [32] | Chowdhury JU, Nandi NC, Uddin M, et al. (2007) Chemical constituents of essential oils from two types of spearmint (Mentha spicata L. and M. cardiaca L.) introduced in Bangladesh. Bangl J Sci Ind Res 42: 79–82. |
| [33] |
Smith SE, Smith FA (2011) Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales. Ann Rev Plant Biol 62: 227–250. doi: 10.1146/annurev-arplant-042110-103846
|
| [34] |
Maya MA, Matsubara Y (2013) Tolerance to Fusarium wilt and anthracnose diseases and changes of antioxidative activity in mycorrhizal cyclamen. Crop Prot 47: 41–48. doi: 10.1016/j.cropro.2013.01.007
|
| [35] | Yang H, Dai Y, Wang X, et al. (2014) Meta-analysis of interactions between arbuscular mycorrhizal fungi and biotic stressors of plants. Sci World J 16: 746506. |
| [36] |
Vokou D, Margaris N, Lynch J (1984) Effects of volatile oils from aromatic shrubs on soil microorganisms. Soil Biol Biochem 16: 509–513. doi: 10.1016/0038-0717(84)90060-9
|
| [37] |
Koske RE, Gemma JN (1989) A modified procedure for staining roots to detect VA mycorrhizas. Mycol Res 92: 486–488. doi: 10.1016/S0953-7562(89)80195-9
|
| [38] |
Orfanoudakis M, Wheeler CT, Hooker JE (2010) Both the arbuscular mycorrhizal fungus Gigaspora rosea and Frankia increase root system branching and reduce root hair frequency in Alnus glutinosa. Mycorrhiza 20: 117–126. doi: 10.1007/s00572-009-0271-0
|
| [39] | Trouvelot A, Kough J, Gianinazzi-Pearson V (1986) Mesure du taux de mycorrhization d'un systeme radiculaire recherché de methods d'estimation ayant une signification fonctionnelle, In: Gininazzi-Pearson V, Giainazzi S, Physiological and genetical aspects of mycorrhiza, Paris: INRA Publications, 217–221. |
| [40] |
Allison SD, Jastrow JD (2006) Activities of extracellular enzymes in physically isolated fractions of restored grassland soils. Soil Biol Biochem 38: 3245–3256. doi: 10.1016/j.soilbio.2006.04.011
|
| [41] |
Sinsabaugh RL, Reynolds H, Long TM (2000) Rapid assay for amidohydrolase (urease) activity in environmental samples. Soil Biol Biochem 32: 2095–2097. doi: 10.1016/S0038-0717(00)00102-4
|
| [42] | Tabatabai M (1994) Soil enzymes, In: Weaver R, Angles J, Bottomley P, Methods of Soil Analysis Part 2, Microbiological and Biochemical Properties, Madison: Soil Science Society of America, 775–833. |
| [43] |
Acosta-Martínez V, Tabatabai MA (2000) Arylamidase activity of soils. Soil Sci Soc Am J 64: 215. doi: 10.2136/sssaj2000.641215x
|
| [44] | Papadopoulou ES, Karpouzas DG, Menkissoglu-Spiroudi U (2011) Extraction parameters significantly influence the quantity and the profile of PLFAs extracted from soils. Microb Ecol 6: 704–714. |
| [45] |
Spyrou IM, Karpouzas DG, Menkissoglu-Spiroudi U (2009) Do botanical pesticides alter the structure of the soil microbial community? Microb Ecol 58: 715–727. doi: 10.1007/s00248-009-9522-z
|
| [46] |
McKinley VL, Peacock AD, White DC (2005) Microbial community PLFA and PHB responses to ecosystem restoration in tallgrass prairie soils. Soil Biol Biochem 37: 1946–1958. doi: 10.1016/j.soilbio.2005.02.033
|
| [47] |
Myers RT, Zak DR, White DC, et al. (2001) Landscape-level patterns of microbial community composition and substrate use in upland forest ecosystems. Soil Sci Soc Am J 65: 359. doi: 10.2136/sssaj2001.652359x
|
| [48] |
Zak DR, Ringelberg DB, Pregitzer KS, et al. (1996) Soil microbial communities beneath Populus grandidentata crown under elevated atmospheric CO2. Ecol Appl 6: 257–262. doi: 10.2307/2269568
|
| [49] |
Rillig MC, Mummey DL, Ramsey PW, et al. (2006) Phylogeny of arbuscular mycorrhizal fungi predicts community composition of symbiosis-associated bacteria. FEMS Microbiol Ecol 57: 389–395. doi: 10.1111/j.1574-6941.2006.00129.x
|
| [50] | Frostegård A, Tunlid A, Bååth E (1993) Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types experimentally exposed to different heavy metals. Appl Environ Microb 59: 3605–3617. |
| [51] |
White D, Stair J, Ringelberg D (1996) Quantitative comparisons of in situ microbial biodiversity by signature biomarker analysis. J Ind Microbiol Biot 17: 185–196. doi: 10.1007/BF01574692
|
| [52] |
Smith GA, Nickels JS, Kerger BD, et al. (1986) Quantitative characterization of microbial biomass and community structure in subsurface material: a prokaryotic consortium responsive to organic contamination. Can J Microbiol 32: 104–111. doi: 10.1139/m86-022
|
| [53] |
Burrows R, Ahmed I (2007) Fungicide seed treatments minimally affect arbuscular-mycorrhizal fungal (AMF) colonization of selected vegetable crops. J Biol Sci 7: 417–420. doi: 10.3923/jbs.2007.417.420
|
| [54] | Huang JC, Lai WA, Singh S, et al. (2013) Response of mycorrhizal hybrid tomato cultivars under saline stress. J Soil Sci Plant Nutr 13: 469–484. |
| [55] |
Nogueira MA, Cardoso EJBN (2007) Phosphorus availability changes the internal and external endomycorrhizal colonization and affects symbiotic effectiveness. Sci Agr 64: 295–300. doi: 10.1590/S0103-90162007000300013
|
| [56] |
Zsögön A, Lambais MR, Benedito VA, et al. (2008) Reduced arbuscular mycorrhizal colonization in tomato ethylene mutants. Sci Agr 65: 259–267. doi: 10.1590/S0103-90162008000300006
|
| [57] | Ortas I, Razzaghi S, Rafique M (2016) Arbuscular mycorrhizae: Effect of rhizosphere and relation with carbon nutrition, In: Choudhary DK, Varma A, Tuteja N, Plant-microbe interaction: An approach to sustainable agriculture, Springer Nature Singapore Pte Ltd, 125–154. |
| [58] |
Baar J, Paradi I, Lucasen EC, et al. (2011) Molecular analysis of AMF diversity in aquatic macrophytes: a comparison of oligotrophic and ultra-oligotrophic lakes. Aquat Bot 94: 53–61. doi: 10.1016/j.aquabot.2010.09.006
|
| [59] | Christensen H, Jakobsen I (1993) Reduction of bacterial growth by a vesicular-arbuscular mycorrhizal fungus in the rhizosphere of cucumber (Cucumis sativus L.). Biol Fertil Soils 1: 253–258. |
| [60] | Zarea M, Ghalavand A, Goltapeh M, et al. (2009) Role of clover species and AM Fungi (Glomus mosseae) on forage yield, nutrients uptake, nitrogenase activity and soil microbial biomass. J Agr Tech 5: 337–347. |
| [61] |
Ladygina N, Henry F, Kant MR, et al. (2010) Additive and interactive effects of functionally dissimilar soil organisms on a grassland plant community. Soil Biol Biochem 42: 2266–2275. doi: 10.1016/j.soilbio.2010.08.027
|
| [62] |
Finlay RD (2008) Ecological aspects of mycorrhizal symbiosis: with special emphasis on the functional diversity of interactions involving the extraradical mycelium. J Exp Bot 59: 1115–1126. doi: 10.1093/jxb/ern059
|
| [63] |
Acosta-Martínez V, Cruz L, Sotomayor-Ramírez D, et al. (2007) Enzyme activities as affected by soil properties and land use in a tropical watershed. Appl Soil Ecol 35: 35–45. doi: 10.1016/j.apsoil.2006.05.012
|
| [64] |
Vierheilig H, Alt M, Mohr U, et al. (1994) Ethylene biosynthesis and activities of chitinase and β-1,3-glucanase in the roots of host and non-host plants of vesicular-arbuscular mycorrhizal fungi after inoculation with Glomus mosseae. J Plant Physiol 143: 337–343. doi: 10.1016/S0176-1617(11)81641-X
|
| [65] |
Wang Y, Lin XG, Yin R, et al. (2006) Effects of arbuscular mycorrhizal inoculation on the growth of Elsholtzia splendens and Zea mays and the activities of phosphatase and urease in a multi-metal-contaminated soil under unsterilized conditions. Appl Soil Ecol 31: 110–119. doi: 10.1016/j.apsoil.2005.03.002
|
| [66] |
Huang H, Zhang S, Wu N, et al. (2009) Influence of Glomus etunicatum/Zea mays mycorrhiza on atrazine degradation, soil phosphatase and dehydrogenase activities, and soil microbial community structure. Soil Biol Biochem 41: 726–734. doi: 10.1016/j.soilbio.2009.01.009
|
| [67] |
Qian K, Wang L, Yin N (2012) Effects of AMF on soil enzyme activity and carbon sequestration capacity in reclaimed mine soil. Int J Min Sci Technol 22: 553–557. doi: 10.1016/j.ijmst.2012.01.019
|
| [68] |
Jin H, Liu J, Liu J, et al. (2012) Forms of nitrogen uptake, translocation, and transfer via arbuscular mycorrhizal fungi: A review. Sci China Life Sci 55: 474–482. doi: 10.1007/s11427-012-4330-y
|
| [69] |
Karamanoli K, Thalassinos G, Karpouzas D, et al. (2012) Are leaf glandular trichomes of oregano hospitable habitats for bacterial growth? J Chem Ecol 38: 476–485. doi: 10.1007/s10886-012-0117-7
|
| [70] |
Köllner EK, Carstens D, Keller E, et al. (2012) Bacterial chitin hydrolysis in two lakes with contrasting trophic statuses. Appl Environ Microb 78: 695–704. doi: 10.1128/AEM.06330-11
|
| [71] |
Stone MN, Plante AF, Casper BB (2013) Plant and nutrient controls on microbial functional characteristics in a tropical oxisol. Plant Soil 373: 893–905. doi: 10.1007/s11104-013-1840-8
|
| [72] | Stainer R, Adelberg E, Ingraham J (1977) General Microbiology, London: Macmillan. |
| [73] | Cox SD, Mann CM, Markham JL, et al. (2000) The mode of antimicrobial action of the essential oil of Melaleuca alternifolia (tea tree oil). J Appl Microbiol 88: 170–175. |
| [74] |
Prashar A, Hili P, Veness RG, et al. (2003) Antimicrobial action of palmarosa oil (Cymbopogon martinii) on Saccharomyces cerevisiae. Phytochemistry 63: 569–575. doi: 10.1016/S0031-9422(03)00226-7
|
| [75] |
Bach EM, Baera SG, Meyera CK, et al. (2010) Soil texture affects soil microbial and structural recovery during grassland restoration. Soil Biol Biochem 42: 2182–2191. doi: 10.1016/j.soilbio.2010.08.014
|
| [76] |
Averill C, Turner BL, Finzi AC (2014) Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature 505: 543–545. doi: 10.1038/nature12901
|
| [77] |
Kaiser C, Koranda M, Kitzler B, et al. (2010) Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol 187: 843–858. doi: 10.1111/j.1469-8137.2010.03321.x
|
| [78] |
Cusack DF, Silver WL, Torn MS, et al. (2011) Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology 92: 621–632. doi: 10.1890/10-0459.1
|
| [79] |
Fragoeiro S, Magan N (2005) Enzymatic activity, osmotic stress and degradation of pesticide mixtures in soil extract liquid broth inoculated with Phanerochaete chrysosporium and Trametes versicolor. Environ Microbiol 7: 348–355. doi: 10.1111/j.1462-2920.2005.00699.x
|
| [80] | Nannipieri P, Giagnoni L, Landi L, et al. (2011) Role of phosphatase enzymes in soil, In: Bünemann EK, Oberson A, Frossard E, Phosphorus in Action, Berlin Heidelberg: Springer, 215–243. |
| [81] | Závodská L, Lesn'y J (2006) Recent development in lignite investigation. HEJ Manuscript: 1418–7108. |
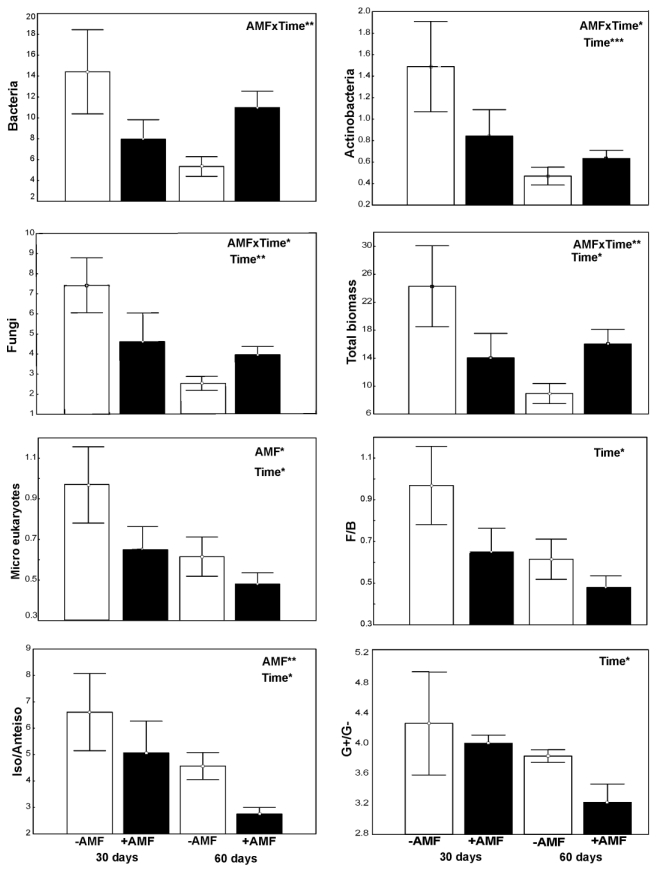
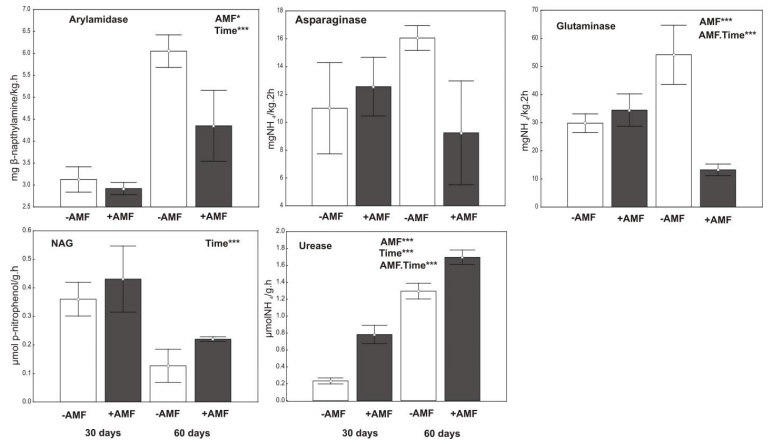
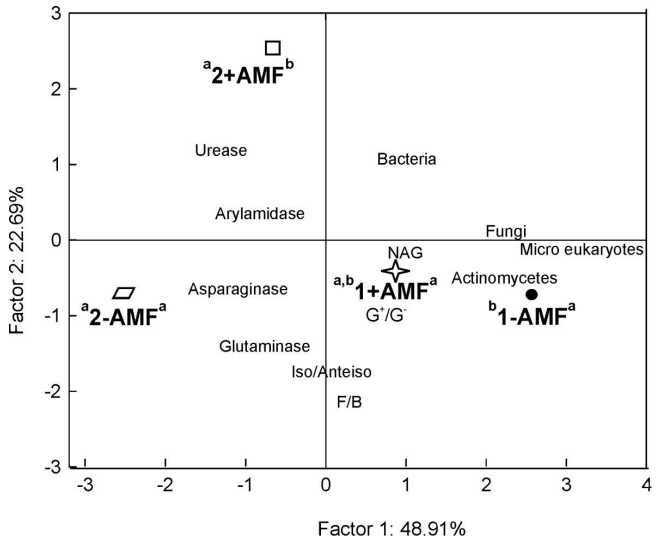
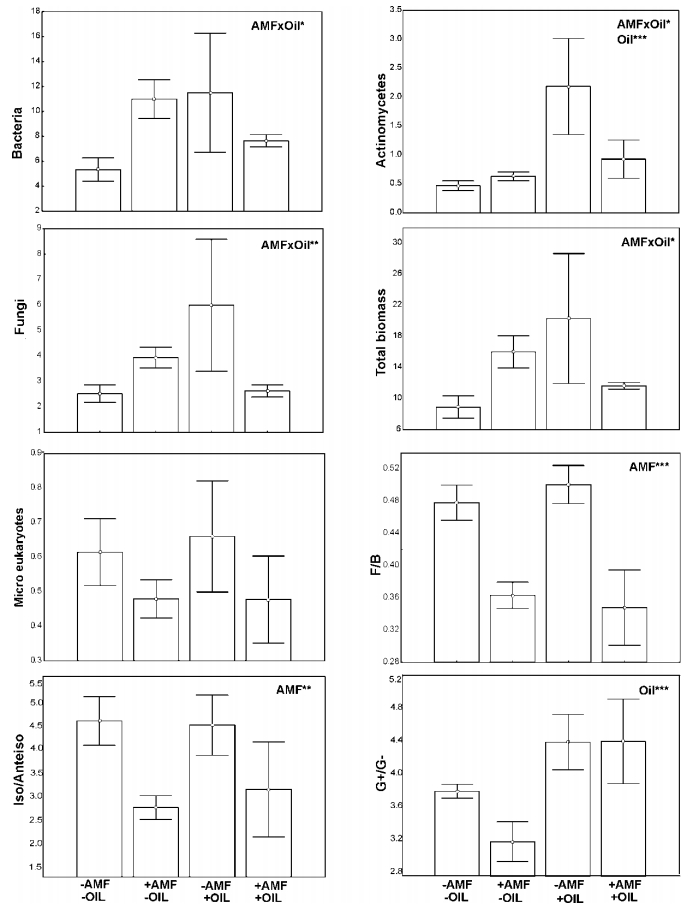
Figures(7) / Tables(1)

George P. Stamou, Sotiris Konstadinou, Nikolaos Monokrousos, Anna Mastrogianni, Michalis Orfanoudakis, Christos Hassiotis, Urania Menkissoglu-Spiroudi, Despoina Vokou, Efimia M. Papatheodorou. The effects of arbuscular mycorrhizal fungi and essential oil on soil microbial community and N-related enzymes during the fungal early colonization phase[J]. AIMS Microbiology, 2017, 3(4): 938-959. doi: 10.3934/microbiol.2017.4.938







 DownLoad:
DownLoad: