Citation: Alexandre A. de Castro, Letícia C. Assis, Daniela R. Silva, Silviana Corrêa, Tamiris M. Assis, Giovanna C. Gajo, Flávia V. Soares, Teodorico C. Ramalho. Computational enzymology for degradation of chemical warfare agents: promising technologies for remediation processes[J]. AIMS Microbiology, 2017, 3(1): 108-135. doi: 10.3934/microbiol.2017.1.108

| [1] | Castro AA, Caetano MS, Silva TC, et al. (2016) Molecular docking, metal substitution and hydrolysis reaction of chiral substrates of phosphotriesterase. Comb Chem High Throughput Screen 19: 334–344. |
| [2] | Calvert GM, Karnik J, Mehler L, et al. (2008) Acute pesticide poisoning among agricultural workers in the United States, 1998–2005. Am J Ind Med 51: 883–898. |
| [3] | Berny P (2007) Pesticides and the intoxication of wild animals. J Vet Pharmacol Ther 30: 93–100. |
| [4] | Lima WEA, Pereira AF, Castro AA, et al. (2016) Flexibility in the molecular design of Acetylcholinesterase Reactivators: Probing Representative Conformations by Chemometric Techniques and Docking/QM Calculations. Lett Drug Des Discov 13: 360–371. |
| [5] | Sartorelli J, Castro AA, Ramalho TC, et al. (2016) Asymmetric biocatalysis of the nerve agent VX by human serum paraoxonase 1: molecular docking and reaction mechanism calculations. Med Chem Res 25: 2521–2533. |
| [6] | King, AM, Aaron CK (2015) Organophosphate and carbamate poisoning. Emerg Med Clin North Am 33: 133–151. |
| [7] | Ramalho TC, Castro AA, Silva DR, et al. (2016) Computational enzymology and organophosphorus degrading enzymes: Promising approaches toward remediation technologies of warfare agents and pesticides. Curr Med Chem 23: 1041–1061. |
| [8] | Iyer R, Iken B, Damania A (2013) A comparison of organophosphate degradation genes and bioremediation applications. Environ Microbiol Rep 5: 787–798. |
| [9] | Iyer R, Iken B (2015) Protein engineering of representative hydrolytic enzymes for remediation of organophosphates. Biochem Eng J 94: 134–144. |
| [10] | Kuca K, Bartosova L, Jun D, et al. (2005) New quaternary pyridine aldoximes as casual antidotes against nerve agents intoxications. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub 149: 75–82. |
| [11] | Nemukhin AV, Grigorenko BL, Lushchekina SV, et al. (2012) Quantum chemical modelling in the research of molecular mechanisms of enzymatic catalysis. Russ Chem Rev 81: 1011–1025. |
| [12] | van der Kamp MW, Mulholland AJ (2008) Computational enzymology: insight into biological catalysts from modelling. Nat Prod Rep 25: 1001–1014. |
| [13] | Puri KD, Di Paolo JA, Gold MR (2013) B-cell receptor signaling inhibitors for treatment of autoimmune inflammatory diseases and B-cell malignancies. Int Rev Immunol 32: 397–427. |
| [14] | Chen YC (2015) Beware of docking! Trends Pharmacol Sci 36: 78–95. |
| [15] | Thomsen R, Christensen MH (2006) MolDock: a new technique for high-accuracy molecular docking. J Med Chem 49: 3315–3321. |
| [16] | Jones G, Willett P, Glen RC, et al. (1997) Development and validation of a genetic algorithm for flexible docking. J Mol Biol 267: 727–748. |
| [17] | Goodsell DS, Olson AJ (1990) Automated docking of substrates to proteins by simulated annealing. Proteins Struct Funct Bioinforma 8: 195–202. |
| [18] | Morris GM, Goodsell DS, Olson AJ (1996) Automated docking of flexible ligands: applications of AutoDock. J Comput Aided Mol Des 10: 293–304. |
| [19] | Rarey M, Kramer B, Lengauer T, et al. (1996) A fast flexible docking method using an incremental construction algorithm. J Mol Biol 261: 470–489. |
| [20] | Azevedo LS, Moraes FP, Xavier MM, et al. (2012) Recent progress of molecular docking simulations applied to development of drugs. Curr Bioinform 7: 352–365. |
| [21] | Liu M, Wang S (1999) MCDOCK: a Monte Carlo simulation approach to the molecular docking problem. J Comput Aided Mol Des 13: 435–451. |
| [22] | Rosenfeld RJ, Goodsell DS, Musah RA, et al. (2003) Automated docking of ligands to an arti cial active site: augmenting crystallographic analysis with computer modeling. J Comput Aided Mol Des 17: 525–536. |
| [23] | da Cunha EFF, Ramalho TC, Reynolds RC (2008) Binding mode analysis of 2,4-diamino-5-methyl-5-deaza-6-substituted pteridines with Mycobacterium tuberculosis and human dihydrofolate reductases. J Biomol Struct Dyn 25: 377–385. |
| [24] | Assis LC, Santos-Garcia L, Ramalho TC, et al. (2013) Interactions of pyrimidine derivatives with dihydrofolate reductase and thymidylate synthase: directions toward combating toxoplasmosis. Curr Bioact Compd 9: 153–166. |
| [25] | Kuntz ID, Blaney JM, Oatley SJ, et al. (1982) A geometric approach to macromolecule-ligand interactions. J Mol Biol 161: 269–288. |
| [26] | Kirkpatrick S, Gelatt Jr CD, Vecchi MP (1983) Optimization by simmulated annealing. Science 220: 671–680. |
| [27] | Assis TM, Mancini DT, Ramalho TC, et al. (2014) In silico study of Leishmania donovani α-β tubulin and inhibitors. J Chem 2014: 1–8. |
| [28] | Giacoppo JOS, Mancini DT, Guimaraes AP, et al. (2015) Molecular modeling toward selective inhibitors of dihydrofolate reductase from the biological warfare agent Bacillus anthracis. Eur J Med Chem 91: 63–71. |
| [29] | Mulholland AJ (2007) Chemical accuracy in QM/MM calculations on enzyme-catalysed reactions. Chem Cent J 1: 1–5. |
| [30] | Santos-Garcia L, Assis LC, Silva DR, et al. (2016) QSAR analysis of nicotinamidic compounds and design of potential Bruton's tyrosine kinase (Btk) inhibitors. J Biomol Struct Dyn 34: 1421–1440. |
| [31] | Namba AM, Silva VB, Silva CHTP (2008) Dinâmica molecular: teoria e aplicações em planejamento de fármacos. Ecl Quim 33: 13–23. |
| [32] | Fattebert JL, Lau EY, Bennion BJ, et al. (2015) Large-scale first-principles molecular dynamics simulations with electrostatic embedding: application to acetylcholinesterase catalysis. J Chem Theory Comput 11: 5688–5695. |
| [33] | de Azevedo WF (2011) Molecular dynamics simulations of protein targets identified in Mycobacterium tuberculosis. Curr Med Chem 18: 1353–1366. |
| [34] | Weiner PK, Kollman PA (1981) AMBER: Assisted model building with energy refinement. A general program for modeling molecules and their interactions. J Comput Chem 2: 287–303. |
| [35] | Brooks BR, Bruccoleri RE, Olafson BD, et al. (1983) CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J Comput Chem 4: 187–217. |
| [36] | Kalé L, Skeel R, Bhandarkar M, et al. (1999) NAMD2: greater scalability for parallel molecular dynamics. J Comput Phys 151: 283–312. |
| [37] | Phillips JC, Braun R, Wang W, et al. (2005) Scalable molecular dynamics with NAMD. J Comput Chem 26: 1781–1802. |
| [38] | Scott WRP, Hünenberger PH, Tironi IG, et al. (1999) The GROMOS biomolecular simulation program package. J Phys Chem A 103: 3596–3607. |
| [39] | Dubey KD, Tiwari RK, Ojha RP (2013) Recent advances in protein-ligand interactions: molecular dynamics simulations and binding free energy. Curr Comput Aided-Drug Des 9: 518–531. |
| [40] | Durrant JD, McCammon JA (2011) Molecular dynamics simulations and drug discovery. BMC Biol 9: 1–9. |
| [41] | Karplus M, McCammon JA (2002) Molecular dynamics simulations of biomolecules. Nat Struct Biol 9: 646–652. |
| [42] | Ochoa R, Rodriguez CA, Zuluaga AF (2016) Perspectives for the structure-based design of acetylcholinesterase reactivators. J Mol Graph Model 68: 176–183. |
| [43] | Zheng M, Waller MP (2016) Adaptive quantum mechanics/molecular mechanics methods. Wiley Interdiscip Rev Comput Mol Sci 6: 369–385. |
| [44] | Ryde, U (2016) QM/MM calculations on proteins, In: Voth GA, Methods in enzymology: computational approaches for studying enzyme mechanism Part A, 1 Ed., Elsevier Inc., 119–158. |
| [45] | Lodola A, Mulholland AJ (2013) Computational enzymology. Methods Mol Biol 924: 67–89. |
| [46] | Senn HM, Thiel W (2009) QM/MM methods for biomolecular systems. Angew Chem Int Ed Engl 48: 1198–1229. |
| [47] | van der Kamp MW, Mulholland AJ (2013) Combined quantum mechanics/molecular mechanics (QM/MM) methods in computational enzymology. Biochemistry 52: 2708–2728. |
| [48] | Silva MC, Torres JA, Castro AA, et al. (2015) Combined experimental and theoretical study on the removal of pollutant compounds by peroxidases: affinity and reactivity toward a bioremediation catalyst. J Biomol Struct Dyn 34: 1–35. |
| [49] | Silva TC, Pires MS, Castro AA, et al. (2015) Molecular insight into the inhibition mechanism of plant and rat 4-hydroxyphenylpyruvate dioxygenase by molecular docking and DFT calculations. Med Chem Res 24: 3958–3971. |
| [50] | Balali-Mood M, Abdollahi M (2014) Basic and clinical toxicology of organophosphorus compounds, 1 Ed., London: Springer International Publishing. |
| [51] | Kumar DN, Alex SA, Chandrasekaran N, et al. (2016) Acetylcholinesterase (AChE)-mediated immobilization of silver nanoparticles for the detection of organophosphorus pesticides. RSC Adv 6: 64769–64777. |
| [52] | Jang YJ, Kim K, Tsay O, et al. (2015) Update 1 of: destruction and detection of chemical warfare agents. Chem Rev 115: PR1–PR76. |
| [53] | Kim K, Tsay OG, Atwood DA, et al. (2011) Destruction and detection of chemical warfare agents. Chem Rev 111: 5345–5403. |
| [54] | Roberts JR, Reigart JR (2013) Recognition and management of pesticide poisoning, 6 Eds., Washington: United States Environmental Protection Agency Office of Pesticide Programs. |
| [55] | Cavalcanti LPAN, Aguiar AP, Lima JA, Lima ALS (2016) Organophosphorous poisoning: treatment and analytical methodologies applied in evaluation of reactivation and inhibition of acetylcholinesterase. Rev Virtual Química 8: 739–766. |
| [56] | Giacoppo JOS, França TCC, Kuča K, et al. (2014) Molecular modeling and in vitro reactivation study between the oxime BI-6 and acetylcholinesterase inhibited by different nerve agents. J Biomol Struct Dyn 33: 2048–2058. |
| [57] | Dvir H, Silman I, Harel M, et al. (2010) Acetylcholinesterase: from 3D structure to function. Chem Biol Interact 187: 10–22. |
| [58] | Patočka J, Cabal J, Kuča, K, et al. (2005) Oxime reactivation of acetylcholinesterase inhibited by toxic phosphorus esters: In vitro kinetics and thermodynamics. J Appl Biomed 3: 91–99. |
| [59] | Tafuri J, Roberts J (1987) Organophosphate poisoning. Ann Emerg Med 16: 193–202. |
| [60] | Mileson B (1998) Common mechanism of toxicity: a case study of organophosphorus pesticides. Toxicol Sci 41: 8–20. |
| [61] | Eddleston M (2004) Self poisoning with pesticides. BMJ 328: 42–44. |
| [62] | Delfino RT, Ribeiro TS, Figueroa-Villar JD (2009) Organophosphorus compounds as chemical warfare agents: A review. J Braz Chem Soc 20: 407–428. |
| [63] | Hörnberg A, Tunemalm AK, Ekström F (2007) Crystal structures of acetylcholinesterase in complex with organophosphorus compounds suggest that the acyl pocket modulates the aging reaction by precluding the formation of the trigonal bipyramidal transition state. Biochemistry 46: 4815–4825. |
| [64] | DuBois, KP (1971) The toxicity of organophosphorus compounds to mammals. Bull World Health Organ 44: 231–240. |
| [65] | Pasquet J, Mazuret A, Fournel J, et al. (1976) Acute oral and percutaneous toxicity of phosalone in the rat, in comparison with azinphosmethyl and Parathion. Toxicol Appl Pharm 37: 85–92. |
| [66] | Jamal GA, Hansen S, Julu PO (2002) Low level exposures to organophosphorus esters may cause neurotoxicity. Toxicology 182: 23–33. |
| [67] | Brunton LL, Chabner BA, Knollmann BC (2010) The Pharmacological Basis of Therapeutics. |
| [68] | Kavalci C, Durukan P, Ozer M, et al. (2009) Organophosphate poisoning due to a wheat bagel. Intern Med 48: 85–88. |
| [69] | Mercey G, Verdelet T, Renou J, et al. (2012) Reactivators of acetylcholinesterase inhibited by organophosphorus nerve agents. Acc Chem Res 45: 756–766. |
| [70] | Wilson IB, Ginsburg S (1955) A powerful reactivator of alkylphosphate-inhibited acetylcholinesterase. Biochim Biophys Acta 18: 168–170. |
| [71] | Soares S, Vieira AA, Delfino RT, et al. (2013) NMR determination of Electrophorus electricus acetylcholinesterase inhibition and reactivation by neutral oximes. Bioorg Med Chem 21: 5923–5930. |
| [72] | Kuca K, Jun D, Musilek K (2006) Structural requirements of acetylcholinesterase reactivators. Mini-Reviews Med Chem 6: 269–277. |
| [73] | Musilek K, Dolezal M, Gunn-Moore F, et al. (2011) Design, evaluation and structure-activity relationship studies of the AChE reactivators against organophosphorus pesticides. Med Res Rev 31: 548–575. |
| [74] | Husain K (2013) Delayed neurotoxicity of organophosphorus compounds. J Enviromental Immunol Toxicol 1: 14–21. |
| [75] | Sahu AK, Sharma R, Gupta B, et al. (2016) Oxime-mediated in vitro reactivation kinetic analysis of organophosphates-inhibited human and electric eel acetylcholinesterase. Toxicol Mech Methods 6516: 1–8. |
| [76] | Szinicz L (2005) History of chemical and biological warfare agents. Toxicology 214: 167–181. |
| [77] | Singh BK, Walker A (2006) Microbial degradation of organophosphorus compounds. FEMS Microbiol Rev 30: 428–471. |
| [78] | Omburo GA, Kuo JM, Mullins LS, et al. (1992) Characterization of the zinc-binding site of bacterial phosphotriesterase. J Biol Chem 267: 13278–13283. |
| [79] | Perezgasga L, Sanchez-Sanchez L, Aguila S, et al. (2012) Substitution of the catalytic metal and protein PEGylation enhances activity and stability of bacterial phosphotriesterase. Appl Biochem Biotechnol 166: 1236–1247. |
| [80] | Tsai PC, Fox N, Bigley AN, et al. (2012) Enzymes for the homeland defense: optimizing phosphotriesterase for the hydrolysis of organophosphate nerve agents. Biochemistry 51: 6463–6475. |
| [81] | Singh BK (2009) Organophosphorus-degrading bacteria: ecology and industrial applications. Nat Rev Microbiol 7: 156–164. |
| [82] | Theriot CM, Grunden AM (2011) Hydrolysis of organophosphorus compounds by microbial enzymes. Appl Microbiol Biotechnol 89: 35–43. |
| [83] | Bigley AN, Raushel FM (2013) Catalytic mechanisms for phosphotriesterases. Biochim Biophys Acta 1834: 443–453. |
| [84] | Rochu D, Viguié N, Renault F, et al. (2004) Contribution of the active-site metal cation to the catalytic activity and to the conformational stability of phosphotriesterase: temperature- and pH-dependence. Biochem J 380: 627–633. |
| [85] | Chen SL, Fang WH, Himo F (2008) Technical aspects of quantum chemical modeling of enzymatic reactions: the case of phosphotriesterase. Theor Chem Acc 120: 515–522. |
| [86] | Vanhooke JL, Benning MM, Raushel FM, et al. (1996) Three-dimensional structure of the zinc-containing phosphotriesterase with the bound substrate analog diethyl 4-methylbenzylphosphonate. Biochemistry 35: 6020–6025. |
| [87] | Lotti M (2002) Promotion of organophosphate induced delayed polyneuropathy by certain esterase inhibitors. Toxicology 181: 245–248. |
| [88] | Chen-Goodspeed M, Sogorb MA, Wu F, et al. (2001) Structural determinants of the substrate and stereochemical specificity of phosphotriesterase. Biochemistry 40: 1325–1331. |
| [89] | Chen-Goodspeed M, Sogorb MA, Wu F, et al. Enhancement, relaxation, and reversal of the stereoselectivity for phosphotriesterase by rational evolution of active site residues. Biochemistry 40: 1332–1339. |
| [90] | Nowlan C, Li Y, Hermann JC, et al. (2006) Resolution of chiral phosphate, phosphonate, and phosphinate esters by an enantioselective enzyme library. J Am Chem Soc 128: 15892–15902. |
| [91] | Hong SB, Raushel FM (1999) Stereochemical preferences for chiral substrates by the bacterial phosphotriesterase. Chem Biol Interact 119: 225–234. |
| [92] | Tsai PC, Bigley A, Li Y, et al. (2010) Stereoselective hydrolysis of organophosphate nerve agents by the bacterial phosphotriesterase. Biochemistry 49: 7978–7987. |
| [93] | Lewis VE, Donarski WJ, Wild JR, et al. (1988) Mechanism and stereochemical course at phosphorus of the reaction catalyzed by a bacterial phosphotriesterase. Biochemistry 27: 1591–1597. |
| [94] | Aubert SD, Li YC, Raushel FM (2004) Mechanism for the hydrolysis of organophosphates by the bacterial phosphotriesterase. Biochemistry 43: 5707–5715. |
| [95] | Bigley AN, Xu C, Henderson TJ, et al. (2013) Enzymatic neutralization of the chemical warfare agent VX: evolution of phosphotriesterase for phosphorothiolate hydrolysis. J Am Chem Soc 135: 10426–10432. |
| [96] | Nachon F, Brazzolotto X, Trovaslet M, et al. (2013) Progress in the development of enzyme-based nerve agent bioscavengers. Chem Biol Interact 206: 536–544. |
| [97] | Furlong CE (2008) Paraoxonases: An historical perspective, In: Mackness B, Mackness M, Aviram M, et al., The paraoxonases: their role in disease development and xenobiotic metabolism, Springer, 3–31. |
| [98] | Aharoni A, Gaidukov L, Khersonsky O, et al. (2005) The 'evolvability' of promiscuous protein functions. Nat Genet 37: 73–76. |
| [99] | Khersonsky O, Tawfik DS (2006) The histidine 115-histidine 134 dyad mediates the lactonase activity of mammalian serum paraoxonases. J Biol Chem 281: 7649–7656. |
| [100] | Cannard K (2006) The acute treatment of nerve agent exposure. J Neurol Sci 249: 86–94. |
| [101] | Gaidukov L, Tawfik DS (2007) The development of human sera tests for HDL-bound serum PON1 and its lipolactonase activity. J Lipid Res 48: 1637–1646. |
| [102] | Yeung DT, Lenz DE, Cerasoli DM (2008) Human paraoxonase I: A potential bioscavenger of organophosphorus nerve agents, In: Mackness B, Mackness M, Aviram M, et al., The paraoxonases: their role in disease development and xenobiotic metabolism, Springer, 151–170. |
| [103] | Lenz DE, Yeung D, Smith JR, et al. (2007) Stoichiometric and catalytic scavengers as protection against nerve agent toxicity: A mini review. Toxicology 233: 31–39. |
| [104] | Rochu D, Chabriere E, Masson P (2007) Human paraoxonase: A promising approach for pre-treatment and therapy of organophosphorus poisoning. Toxicology 233: 47–59. |
| [105] | La Du BN, Aviram M, Billecke S, et al. (1999) On the physiological role(s) of the paraoxonases. Chem Biol Interact 119: 379–388. |
| [106] | Ahmed Z, Ravandi A, Maguire GF, et al. (2002) Multiple substrates for paraoxonase-1 during oxidation of phosphatidylcholine by peroxynitrite. Biochem Biophys Res Commun 290: 391–396. |
| [107] | Shih DM, Gu LJ, Xia YR, et al. (1998) Mice lacking serum paraoxonase are susceptible to organophosphate toxicity and atherosclerosis. Nature 394: 284–287. |
| [108] | Mackness B, Mackness M (2010) Anti-inflammatory properties of paraoxonase-1 in atherosclerosis, In: Reddy ST, Paraoxonases in inflammation, infection, and toxicology, Humana Press, 143–151. |
| [109] | Koren-Gluzer M, Aviram M, Meilin E, et al. (2011) The antioxidant HDL-associated paraoxonase-1 (PON1) attenuates diabetes development and stimulates beta-cell insulin release. Atherosclerosis 219: 510–518. |
| [110] | Li WF, Furlong CE, Costa LG (1995) Paraoxonase protects against chlorpyrifos toxicity in mice. Toxicol Lett 76: 219–226. |
| [111] | Camps J, Pujol I, Ballester F, et al. (2011) Paraoxonases as potential antibiofilm agents: their relationship with quorum-sensing signals in gram-negative bacteria. Antimicrob Agents Chemother 55: 1325–1331. |
| [112] | Yeung DT, Lenz DE, Cerasoli DM (2005) Analysis of active-site amino-acid residues of human serum paraoxonase using competitive substrates. FEBS J 272: 2225–2230. |
| [113] | Harel M, Aharoni A, Gaidukov L, et al. (2004) Structure and evolution of the serum paraoxonase family of detoxifying and anti-atherosclerotic enzymes. Nat Struct Mol Biol 11: 412–419. |
| [114] | Fairchild SZ, Peterson MW, Hamza A, et al. (2011) Computational characterization of how the VX nerve agent binds human serum paraoxonase 1. J Mol Model 17: 97–109. |
| [115] | Kuo CL, La Du BN (1998) Calcium binding by human and rabbit serum paraoxonases-structural stability and enzymatic activity. Drug Metab Dispos 26: 653–660. |
| [116] | Blum MM, Timperley CM, Williams GR, et al. (2008) Inhibitory potency against human acetylcholinesterase and enzymatic hydrolysis of Fluorogenic nerve agent mimics by human paraoxonase 1 and squid diisopropyl fluorophosphatase. Biochemistry 47: 5216–5224. |
| [117] | Hu X, Jiang X, Lenz DE, et al. (2009) In silico analyses of substrate interactions with human serum paraoxonase 1. Proteins 75: 486–498. |
| [118] | Otto TC, Harsch CK, Yeung DT, et al. (2009) Dramatic differences in organophosphorus hydrolase activity between human and chimeric recombinant mammalian paraoxonase-1 enzymes. Biochemistry 48: 10416–10422. |
| [119] | Sanan TT, Muthukrishnan S, Beck JM, et al. (2010) Computational modeling of human paraoxonase 1: preparation of protein models, binding studies, and mechanistic insights. J Phys Org Chem 23: 357–369. |
| [120] | Dumas DP, Caldwell SR, Wild JR, et al. (1989) Purification and properties of the phosphotriesterase from Pseudomonas diminuta. J Biol Chem 264: 19659–19665. |
| [121] | Muthukrishnan S, Shete VS, Sanan TT, et al. (2012) Mechanistic insights into the hydrolysis of organophosphorus compounds by paraoxonase-1: exploring the limits of substrate tolerance in a promiscuous enzyme. J Phys Org Chem 25: 1247–1260. |
| [122] | Wymore T, Field MJ, Langan P, et al. (2014) Hydrolysis of DFP and the nerve agent (S)-sarin by DFPase proceeds along two different reaction pathways: Implications for engineering bioscavengers. J Phys Chem B 118: 4479–4489. |
| [123] | Chen JCH, Mustyakimov M, Schoenborn BP, et al. (2010) Neutron structure and mechanistic studies of diisopropyl fluorophosphatase (DFPase). Acta Crystallogr D 66: 1131–1138. |
| [124] | Katsemi V, Lücke C, Koepke J, et al. (2005) Mutational and structural studies of the diisopropylfluorophosphatase from Loligo vulgaris shed new light on the catalytic mechanism of the enzyme. Biochemistry 44: 9022–9033. |
| [125] | Elias M, Liebschner D, Koepke J, et al. (2013) Hydrogen atoms in protein structures: high-resolution X-ray diffraction structure of the DFPase. BMC Res Notes 6: 1–7. |
| [126] | Hoskin FCG, Rosenberg P, Brzin M (1966) Re-examination of the effect of DFP on electrical and cholinesterase activity of squid giant axon. Proc Natl Acad Sci USA 55: 1231–1235. |
| [127] | Hoskin FCG, Long RJ (1972) Purification of a DFP-hydrolyzing enzyme from squid head ganglion. Arch Biochem Biophys 150: 548–555. |
| [128] | Gay DD, Hoskin FCG (1979) Stereospecificity and active site requirements in a diisopropylphosphorofluoridate-hydrolyzing enzyme. Biochem Pharmacol 28: 1259–1261. |
| [129] | Hoskin FCG, Kirkish MA, Steinmann KE (1984) Two enzymes for the detoxication of organophosphorus compounds-sources, similarities, and significance. Fundam Appl Toxicol 4: 165–172. |
| [130] | Hartleib J, Rüterjans H (2001) Insights into the reaction mechanism of the diisopropyl fluorophosphatase from Loligo vulgaris by means of kinetic studies, chemical modification and site-directed mutagenesis. Biochim Biophys Acta 1546: 312–324. |
| [131] | Hartleib J, Geschwindner S, Scharff EI, et al. (2001) Role of calcium ions in the structure and function of the di-isopropylfluorophosphatase from Loligo vulgaris. Biochem J 353: 579–589. |
| [132] | Hartleib J, Rüterjans H (2001) High-yield expression, purification, and characterization of the recombinant diisopropylfluorophosphatase from Loligo vulgaris. Protein Expr Purif 21: 210–219. |
| [133] | Scharff EI, Koepke J, Fritzsch G, et al. (2001) Crystal structure of diisopropylfluorophosphatase from Loligo vulgaris. Structure 9: 493–502. |
| [134] | Blum MM, Löhr F, Richardt A, et al. (2006) Binding of a designed substrate analogue to diisopropyl fluorophosphatase: Implications for the phosphotriesterase mechanism. J Am Chem Soc 128: 12750–12757. |
| [135] | Melzer M, Chen JCH, Heidenreich A, et al. (2009) Reversed enantioselectivity of diisopropyl fluorophosphatase against organophosphorus nerve agents by rational design. J Am Chem Soc 131: 17226–17232. |
| [136] | Koepke J, Scharff EI, Lücke C, et al. (2003) Statistical analysis of crystallographic data obtained from squid ganglion DFPase at 0.85 A resolution. Acta Crystallogr D 59: 1744–1754. |
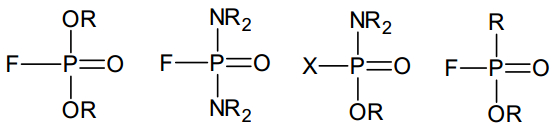
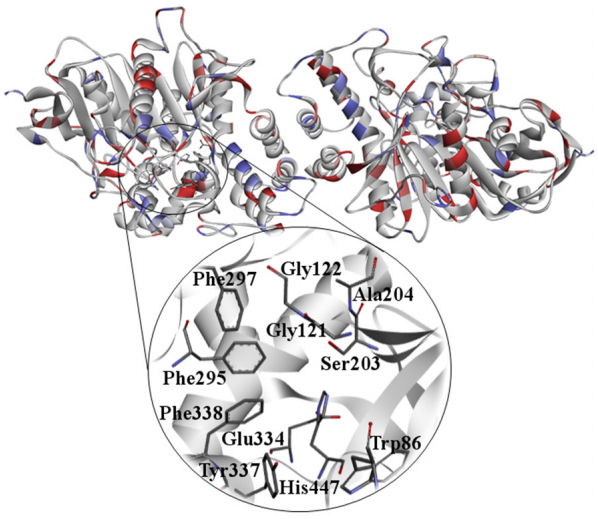
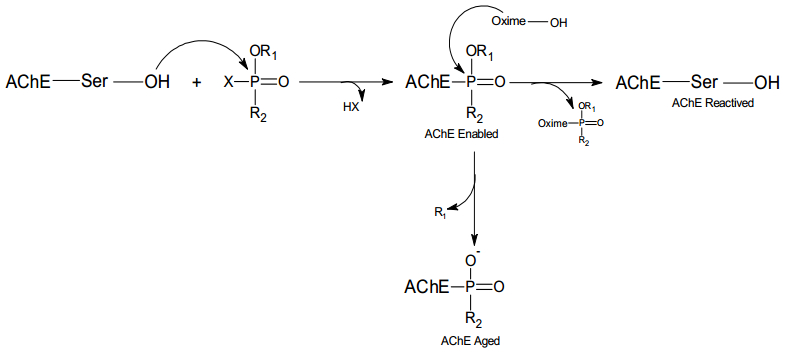
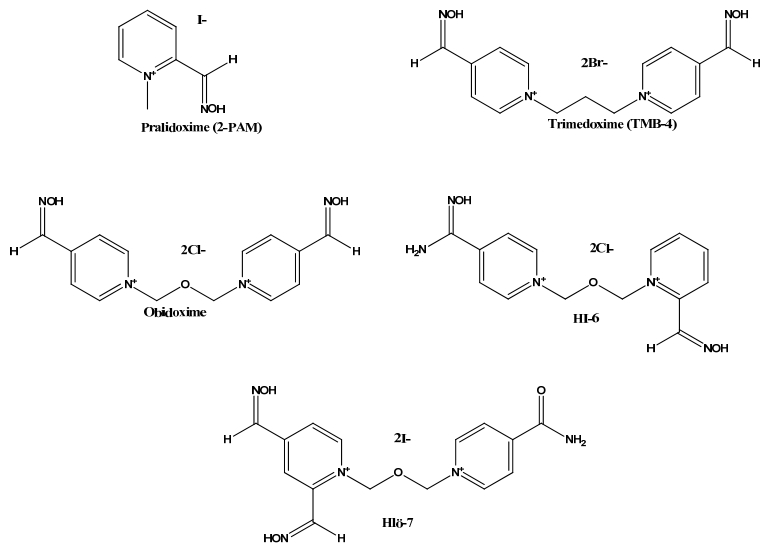
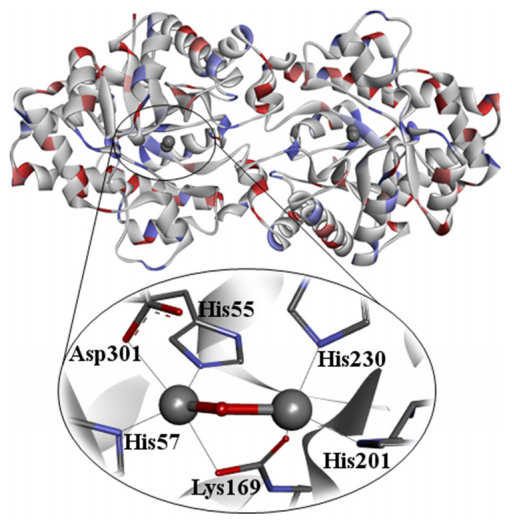
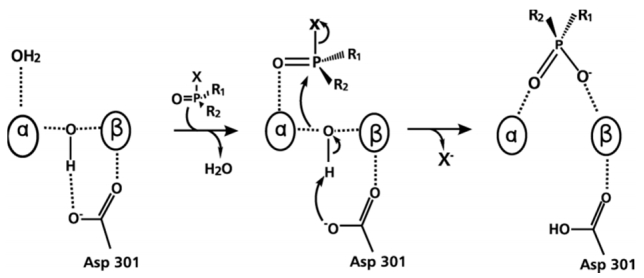
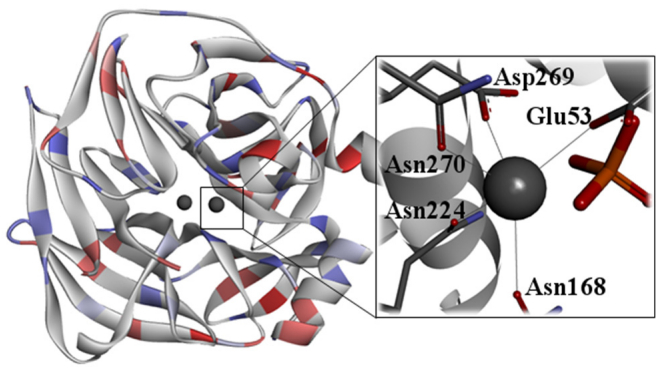
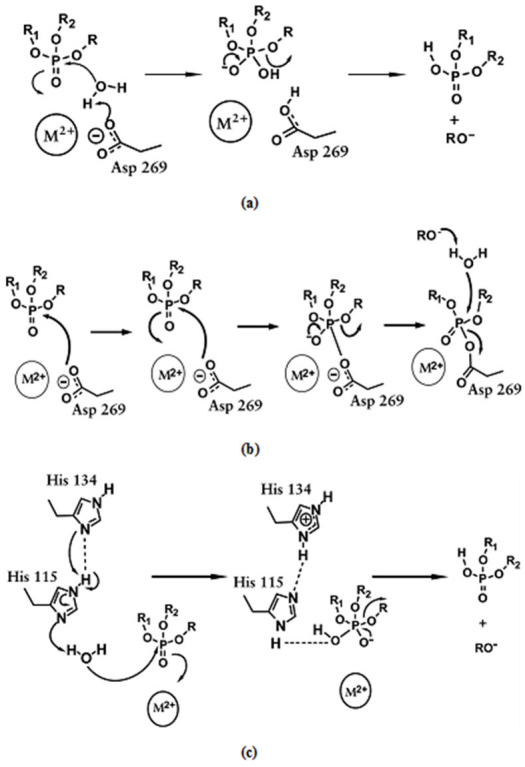
Figures(10)

Alexandre A. de Castro, Letícia C. Assis, Daniela R. Silva, Silviana Corrêa, Tamiris M. Assis, Giovanna C. Gajo, Flávia V. Soares, Teodorico C. Ramalho. Computational enzymology for degradation of chemical warfare agents: promising technologies for remediation processes[J]. AIMS Microbiology, 2017, 3(1): 108-135. doi: 10.3934/microbiol.2017.1.108







 DownLoad:
DownLoad: