Citation: Javier Fernández, Sául Redondo-Blanco, Elisa M. Miguélez, Claudio J. Villar, Alfonso Clemente, Felipe Lombó. Healthy effects of prebiotics and their metabolites against intestinal diseases and colorectal cancer[J]. AIMS Microbiology, 2015, 1(1): 48-71. doi: 10.3934/microbiol.2015.1.48

| [1] |
Eckburg PB, Bik EM, Bernstein CN, et al. (2005) Diversity of the human intestinal microbial flora. Science 308: 1635-1638. doi: 10.1126/science.1110591

|
| [2] |
Qin J, Li R, Raes J, et al. (2010). A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464: 59-65. doi: 10.1038/nature08821
|
| [3] |
Human Microbiome Project Consortium (2012) Structure, function and diversity of the healthy human microbiome. Nature 486: 207-214. doi: 10.1038/nature11234
|
| [4] |
Corthier G, Dore J (2010) A new era in gut research concerning interactions between microbiota in human health. Gastroenterologie Clinique Biologique 34: S1-S6. doi: 10.1016/S0399-8320(10)70014-2
|
| [5] |
Flint HJ, Scott KP, Louis O, et al. (2012) The role of the gut microbiota in nutrition and health. Nat Rev Gastroenterol Hepatol 9: 577-589. doi: 10.1038/nrgastro.2012.156
|
| [6] |
Attene-Ramos MS, Wagner ED, Plewa MJ, et al. (2006) Evidence that hydrogen sulfide is a genotoxic agent. Mol Cancer Res 4: 9-14. doi: 10.1158/1541-7786.MCR-05-0126
|
| [7] |
Nyangale EP, Mottram DS, Gibson GR (2012) Gut microbial activity, implications for health and disease: the potential role of metabolite analysis. J Proteome Res 11: 5573-5585. doi: 10.1021/pr300637d
|
| [8] |
Cerf-Bensussan N, Gaboriau-Routhiau V (2010) The immune system and the gut microbiota: friends or foes? Nat Rev Immunol 10: 735-744. doi: 10.1038/nri2850
|
| [9] |
Sommer F, Backhed F (2013) The gut microbiota-masters of host development and physiology. Nat Rev Microbiol 11: 227-238 doi: 10.1038/nrmicro2974
|
| [10] |
Clemente JC, Ursell LK, Parfrey LW, et al. (2012) The impact of the gut microbiota on human health: an integrative view. Cell 148: 1258-1270. doi: 10.1016/j.cell.2012.01.035
|
| [11] |
Aguirre M, Venema K (2015) The art of targeting gut microbiota for tackling human obesity. Genes Nutr 10: 20. doi: 10.1007/s12263-015-0472-4
|
| [12] |
Hold GL, Smith M, Grange C, et al. (2014) Role of the gut microbiota in inflammatory bowel disease pathogenesis: What have we learnt in the past 10 years? World J Gastroenterol 20: 1192-1210. doi: 10.3748/wjg.v20.i5.1192
|
| [13] |
Wang Z, Klipfell E, Bennett B, et al. (2011) Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 472: 57-65. doi: 10.1038/nature09922
|
| [14] |
Brusca SB, Abramson SB, Scher JU (2014) Microbiome and mucosal inflammation as extra-articular triggers for rheumatoid arthritis and autoimmunity. Curr Opin Rheumatol 26: 101-107. doi: 10.1097/BOR.0000000000000008
|
| [15] | Gao Z, Guo B, Gao R, et al. (2015) Microbiota dysbiosis is associated with colorectal cancer. Front Microbiol 6:20. |
| [16] | Mima K, Nishihara R, Qian ZR, et al. (2015) Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut. pii: gutjnl-2015-310101. doi: 10.1136/gutjnl-2015-310101. |
| [17] | Gibson GR, Roberfroid MB (1995) Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J Nutr 125: 1401-1412. |
| [18] |
Koenig JE, Spor A, Scalfone N, et al. (2011) Succession of microbial consortia in the developing infant gut microbiome. Proc Natl Acad Sci U S A 108: 4578-4585. doi: 10.1073/pnas.1000081107
|
| [19] |
Albenberg LG, Wu GD (2014) Diet and the intestinal microbiome: associations, functions, and implications for health and disease. Gastroenterology 146: 1564-1572. doi: 10.1053/j.gastro.2014.01.058
|
| [20] |
De Filipo C, Cavalieri D, Di Paola M, et al. (2010) Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Nattl Acad Sci U S A 107: 14691-14696. doi: 10.1073/pnas.1005963107
|
| [21] | David LA, Maurice CF, Carmody RN, et al. (2014) Diet rapidly and reproducibly alters the gut microbiome. Nature 505: 559-566. |
| [22] |
Wu GD, Chen J, Hoffmann C, et al. (2011) Linking long-term dietary patterns with gut microbial enterotypes. Science 334: 105-108. doi: 10.1126/science.1208344
|
| [23] |
Simpson HL, Cambell BJ (2015) Review article: Dietary fibre-microbiota interactions. Aliment Pharmocol Ther 42: 158-179. doi: 10.1111/apt.13248
|
| [24] |
Walker AW, Ince J, Duncam SH, et al. (2011) Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J 5: 220-230. doi: 10.1038/ismej.2010.118
|
| [25] | Bouhnik Y, Raskine L, Vicaut E, et al. (2004) The capacity of nondigestible carbohydrates to stimulate fecal bifidobacteria in healthy humans. A double-blind, randomized, placebo-controlled, parallel-group, dose-response relation study. Am J Clin Nutr 80: 1658-1664. |
| [26] |
Saito Y, Hamanaka Y, Saito K, et al. (2002) Stability of species composition of fecal bifidobacteria in human subjects during fermented milk administration. Curr Microbiol 44:368-373. doi: 10.1007/s00284-001-0012-x
|
| [27] |
Muegge BD, Kuczynski J, Knights D, et al. (2011) Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 332: 970-974. doi: 10.1126/science.1198719
|
| [28] |
Sonnenburg ED, Sonnenburg JL (2014) Starving our microbial self: the deleterious consequences of a diet deficient in microbiota-accessible carbohydrates. Cell Metab 20: 779-786. doi: 10.1016/j.cmet.2014.07.003
|
| [29] | Roberfroid M, Gibson GR, Hoyles L, et al. (2010) Prebiotic effects: metabolic and health benefits. Br J Nutr 104: S1-S63. |
| [30] | Gibson GR (2004) From probiotics to prebiotics and a healthy digestive system. J Food Sci 69: 141-143. |
| [31] | Gibson GR, Scott KP, Rastall RA, et al. (2010) Dietary prebiotics: current status and new definition. Food Sci Technol Bull 7: 1-19. |
| [32] |
Bindels LB, Delzenne NM, Cani PD, et al. (2015) Towards a more comprehensive concept for prebiotics. Nat Rev Gastroenterol Hepatol 12: 303-310. doi: 10.1038/nrgastro.2015.47
|
| [33] |
Guarner F, Khan AG, Garish J, et al. (2012) World Gastroenterology Organization Global Guidelines Probiotics and prebiotics. J Clin Gastroenterol 46: 468-481. doi: 10.1097/MCG.0b013e3182549092
|
| [34] |
Bindels LB, Porporato P, Dewulf EM, et al. (2012) Gut microbiota-derived propionate reduces cancer cell proliferation in the liver. Br J Cancer 107: 1337-1344. doi: 10.1038/bjc.2012.409
|
| [35] | Clemente A (2014) In vivo assessment of the bioactivity of food oligosaccharides. In: Moreno FJ, Sanz ML, editors. Wiley Blackwell, Food oligosaccharides: production, analysis and bioactivity; pp 238-254. |
| [36] |
Charalampopoulos D, Rastall RA (2012) Prebiotics in foods. Curr Opin Biotechnol 23: 187-191. doi: 10.1016/j.copbio.2011.12.028
|
| [37] | Corzo N, Alonso JL, Azpiroz F, et al. (2015) Prebiotics: concept, properties and beneficial effects. Nutr Hosp 31: 99-118. |
| [38] |
Rastall RA (2010) Functional oligosaccharides: application and manufacture. Annu Rev Food Sci Technol 1: 305-339. doi: 10.1146/annurev.food.080708.100746
|
| [39] |
Vijn I, Smeekens S (1999) Fructan: more than a reserve carbohydrate? Plant Physiol 120: 351-60. doi: 10.1104/pp.120.2.351
|
| [40] |
Comparot-Moss S, Denyer K (2009) The evolution of the starch biosynthetic pathway in cereals and other grasses. J Exp Bot 60: 2481-2492. doi: 10.1093/jxb/erp141
|
| [41] |
Gallardo K, Thompson R, Burstin J (2008) Reserve accumulation in legume seeds. C R Biol 331: 755-762. doi: 10.1016/j.crvi.2008.07.017
|
| [42] |
Bahaji A, Li J, Sánchez-López ÁM, et al. (2014) Starch biosynthesis, its regulation and biotechnological approaches to improve crop yields. Biotechnol Adv 32: 87-106. doi: 10.1016/j.biotechadv.2013.06.006
|
| [43] |
Ao Z, Simsek S, Zhang G, et al. (2007) Starch with a slow digestion property produced by altering its chain length, branch density, and crystalline structure. J Agric Food Chem 55: 4540-45547. doi: 10.1021/jf063123x
|
| [44] | Galand G (1989) Brush border membrane sucrase-isomaltase, maltase-glucoamylase and trehalase in mammals. Comparative development, effects of glucocorticoids, molecular mechanisms, and phylogenetic implications. Comp Biochem Physiol B 94: 1-11. |
| [45] |
Cairns AJ (2003) Fructan biosynthesis in transgenic plants. J Exp Bot 54: 549-567. doi: 10.1093/jxb/erg056
|
| [46] |
Hernandez-Hernandez O, Marín-Manzano MC, Rubio L, et al. (2012). Monomer and linkage type of galacto-oligosaccharides affect their resistance to ileal digestion and prebiotic properties in rats. J Nutr 142: 1232-1239. doi: 10.3945/jn.111.155762
|
| [47] |
Chi ZM, Zhang T, Cao TS, et al. (2011). Biotechnological potential of inulin for bioprocesses. Bioresour Technol 102: 4295-4303. doi: 10.1016/j.biortech.2010.12.086
|
| [48] |
Ernst MK, Chatterton NJ, Harrison PA, et al. (1998) Characterization of fructan oligomers from species of the genus Allium L. J Plant Physiol 153: 53-60. doi: 10.1016/S0176-1617(98)80044-8
|
| [49] | Roberfroid MB, Van Loo JAE, Gibson GR (1998) The bifidogenic nature of chicory inulin and its hydrolysis products. J Nutrition 128: 11-19. |
| [50] |
Yun JW (1996) Fructooligosaccharides—occurrence, preparation, and application. Enzyme Microbial Technol 19: 107-117. doi: 10.1016/0141-0229(95)00188-3
|
| [51] | Macfarlane GT, Steed H, Macfarlane S (2008) Bacterial metabolism and health-related effects of galacto-oligosaccharides and other prebiotics. J Appl Microbiol 104: 305-344. |
| [52] |
Al-Tamimi MAHM, Palframan RJ, Cooper JM, et al. (2006) In vitro fermentation of sugar beet arabinan and arabinooligosaccharides by the human gut microflora. J Appl Microbiol 100: 407-414. doi: 10.1111/j.1365-2672.2005.02780.x
|
| [53] | Hotchkiss AT, Nunez A, Rastall RA, et al. (2010) Growth promotion of beneficial bacteria in gut of human comprises administering composition comprising arabino oligosaccharide as prebiotic. U.S. patent 2010316766-A1. |
| [54] | Gómez B, Miguez B, Veiga A, et al. (2015) Production, purification and in vitro evaluation of the prebiotic potential of arabinoxylooligosaccharides from breweŕs spent grain. J Agric Food Chem [In press]. |
| [55] |
Palframan RJ, Gibson GR, Rastall RA (2002) Effect of pH and dose on the growth of gut bacteria on prebiotic carbohydrates in vitro. Anaerobe 8: 287-292. doi: 10.1006/anae.2002.0434
|
| [56] |
Rycroft CE, Jones MR, Gibson GR, et al. (2001) A comparative in vitro evaluation of the fermentation properties of prebiotic oligosaccharides. J Appl Microbiol 91: 878-887. doi: 10.1046/j.1365-2672.2001.01446.x
|
| [57] |
Sanz ML, Gibson GR, Rastall RA (2005) Influence of disaccharide structure on prebiotic selectivity in vitro. J Agric Food Chem 53: 5192-5199. doi: 10.1021/jf050276w
|
| [58] |
Sulek K, Vigsnaes LK, Schmidt LR, et al. (2014) A combined metabolomic and phylogenetic study reveals putatively prebiotic effects of high molecular weight arabino-oligosaccharides when assessed by in vitro fermentation in bacterial communities derived from humans. Anaerobe 28: 68-77. doi: 10.1016/j.anaerobe.2014.05.007
|
| [59] |
Westphal Y1, Kühnel S, de Waard P, et al. (2010) Branched arabino-oligosaccharides isolated from sugar beet arabinan. Carbohydr Res 345: 1180-1189. doi: 10.1016/j.carres.2010.03.042
|
| [60] |
Thomassen LV, Vigsnaes LK, Licht TR, et al. (2011) Maximal release of highly bifidogenic soluble dietary fibers from industrial potato pulp by minimal enzymatic treatment. Appl Microbiol Biotechnol 90: 873-884. doi: 10.1007/s00253-011-3092-y
|
| [61] |
Vigsnæs LK, Holck J, Meyer AS, et al. (2011) In Vitro Fermentation of Sugar Beet Arabino-Oligosaccharides by Fecal Microbiota Obtained from Patients with Ulcerative Colitis To Selectively Stimulate the Growth of Bifidobacterium spp. and Lactobacillus spp. Appl Env Microbiol 77: 8336-8344. doi: 10.1128/AEM.05895-11
|
| [62] |
Chung Y, Hsu C, Ko C, et al. (2007). Dietary intake of xylooligosaccharides improves the intestinal microbiota, fecal Moisture, and pH Value in the elderly. Nutr Res 27: 756-761. doi: 10.1016/j.nutres.2007.09.014
|
| [63] |
Lecerf JM, Depeint F, Clerc E, et al. (2012) Xylo-oligosaccharide (XOS) in combination with inulin modulates both the intestinal environment and immune status in healthy subjects, while XOS alone only shows prebiotic properties. Br J Nutr 108: 1847-1858. doi: 10.1017/S0007114511007252
|
| [64] |
Sheu WHH, Lee IT, Chen W, et al. (2008) Effects of xylooligosaccharides in type 2 diabetes mellitus. J Nutr Sci Vitaminol 54: 396-401. doi: 10.3177/jnsv.54.396
|
| [65] | Koga Y, Shibuta T, O'Brien R (1993) Soybean oligosaccharides. In Oligosaccharides. Production, properties and applications. Japan. Technol. Rev. Section E: Biotechnology vol. 3, No 2. Nakakuki T Ed; Gordon and Breach Science Publishers: Switzerland, pp. 175-203 |
| [66] |
Li T, Lu X, Yang X (2013) Stachyose-enriched α-galacto-oligosaccharides regulate gut microbiota and relieve constipation in mice. J Agric Food Chem 61: 11825-11831. doi: 10.1021/jf404160e
|
| [67] |
Ao Z, Quezada-Calvillo R, Sim L, et al. (2007) Evidence of native starch degradation with human small intestinal maltase-glucoamylase (recombinant). FEBS Lett 581: 2381-2388. doi: 10.1016/j.febslet.2007.04.035
|
| [68] | Cardelle-Cobas A, Olano A, Corzo N, et al. (2012) In vitro fermentation of lactulose-derived oligosaccharides by mixed fecal microbiota. J Agric Food Chem 60: 20124-2032. |
| [69] |
Marín-Manzano MC, Abecia L, Hernandez-Hernandez O, et al. (2013) Galacto-oligosaccharides derived from lactulose exert a selective stimulation on the growth of Bifidobacterium animalis in the large intestine of growing rats. J Agric Food Chem 61: 7560-7567. doi: 10.1021/jf402218z
|
| [70] |
Georgi G, Bartke N, Wiens F, et al. (2013) Functional glycans and glycoconjugates in human milk. Am J Clin Nutr 98: 578S-85S. doi: 10.3945/ajcn.112.039065
|
| [71] |
Li M, Bauer LL, Chen X, et al. (2012). Microbial composition and in vitro fermentation patterns of human milk oligosaccharides and prebiotics differ between formula-fed and sow-reared piglets. J Nutr 142: 681-689. doi: 10.3945/jn.111.154427
|
| [72] |
Barile D, Rastall RA (2013) Human milk and related oligosaccharidesas prebiotics. Curr Opin Biotechnol 24: 214-219. doi: 10.1016/j.copbio.2013.01.008
|
| [73] |
Ruiz-Moyano S, Totten SM, Garrido DA, et al. (2013) Variation in consumption of human milk oligosaccharides by infant gut-associated strains of Bifidobacterium breve. Appl Environ Microbiol 79: 6040-6049. doi: 10.1128/AEM.01843-13
|
| [74] | Martin-Ortiz A, Barile D, Salcedo J, et al. (2015) Characterization of goat colostrum oligosaccharides by nano-liquid chromatography on chip quadrupole time of flight mass spectrometry and hydrophilic interaction liquid chromatography-quadrupole mass spectrometry. J Chrom A [in press]. |
| [75] |
Lara-Villoslada F, Debras E, Nieto A, et al. (2006) Oligosaccharides isolated from goat milk reduce intestinal inflammation in a rat model of dextran sodium sulfate-induced colitis. Clin Nutr 25: 477-488. doi: 10.1016/j.clnu.2005.11.004
|
| [76] |
Padilla B, Frau F, Ruiz-Matute AI, et al. (2015). Production of lactulose oligosaccharides by isomerisation of transgalactosylated cheese whey permeate obtained by β-galactosidases from dairy Kluyveromyces. J Dairy Res 82: 356-364. doi: 10.1017/S0022029915000217
|
| [77] |
Panesar PS, Kumari S (2011) Lactulose: Production, purification and potential applications. Biotechnol Adv 29: 940-948. doi: 10.1016/j.biotechadv.2011.08.008
|
| [78] |
Olano A, Corzo N (2009) Lactulose as a food ingredient. J Sci Food Agric 89: 1987-1990. doi: 10.1002/jsfa.3694
|
| [79] | EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA) (2010) Scientific opinion on the substantiation of healthclaims related to lactulose and decreasing potentially pathogenicgastro-intestinal microorganisms (ID 806) and reductionin intestinal transit time (ID 807) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J 8: 1806-1821. |
| [80] | Schumann C (2002) Medical, nutritional and technological properties of lactulose. An update. Eur J Nutr 41:17-25. |
| [81] |
Foster KJ, Lin S, Turck CJ (2010) Current and emerging strategies for treating hepatic encephalopathy. Crit Care Nurs Clin North Am 22: 341-350. doi: 10.1016/j.ccell.2010.04.007
|
| [82] |
Sussman NL (2015) Treatment of Overt Hepatic Encephalopathy. Clin Liver Dis 19: 551-563. doi: 10.1016/j.cld.2015.04.005
|
| [83] | Kaneko T, Yokoyama A, Suzuki M (1995) Digestibility characteristics of isomalto-oligosaccharides in comparison with several saccharides using the rat jejunum loop method. Biosci Biotechnol Biochem 9: 1190-1194. |
| [84] |
Hu Y, Ketabi A, Buchko A, et al. (2013) Metabolism of isomalto-oligosaccharides by Lactobacillus reuteri and bifidobacteria. Lett Appl Microbiol 57: 108-114. doi: 10.1111/lam.12076
|
| [85] |
Ketabi A, Dieleman LA, Gänzle MG (2011) Influence of isomalto-oligosaccharides on intestinal microbiota in rats. J Appl Microbiol 110: 1297-1306. doi: 10.1111/j.1365-2672.2011.04984.x
|
| [86] | Yen CH, Tseng YH, Kuo YW, et al. (2011) Long-term supplementation of isomalto-oligosaccharides improved colonic microflora profile, bowel function, and blood cholesterol levels in constipated elderly people-a placebo-controlled, diet-controlled trial. Nutrition 17: 445-450. |
| [87] | Wang HF, Lim P-S, Kao MD, et al. (2011) Use of isomalto-oligosaccharide in the treatment of lipid profiles and constipation in hemodialysis patients. J Renal Nutr 11: 73-79. |
| [88] | Mizubuchi H, Yajima T, Aoi N, et al. (2005) Isomalto-Oligosaccharides Polarize Th1-Like Responses in Intestinal and Systemic Immunity in Mice. J Nutr 20: 2857-2861. |
| [89] |
Valette P, Pelenc V, Djouzi Z, et al. (1993) Bioavailability of new synthesised glucooligosaccharides in the intestinal tract of gnotobiotic rats. J Sci Food Agricult 62:121-127. doi: 10.1002/jsfa.2740620204
|
| [90] |
Chung CH, Day DF (2002) Glucooligosaccharides from Leuconostoc mesenteroides B-742 (ATCC 13146): a potential prebiotic. J Indus Microbiol Biotechnol 29: 196-199. doi: 10.1038/sj.jim.7000269
|
| [91] |
Sarbini SR, Kolida S, Gibson GR, et al. (2013) In vitro fermentation of commercial α-gluco-oligosaccharide by faecal microbiota from lean and obese human subjects. Bri J Nutr 109: 1980-1989 doi: 10.1017/S0007114512004205
|
| [92] | Wichienchot S, Prasertsan P, Hongpattarakere T, et al. (2006) In vitro Fermentation of Mixed Linkage Glucooligosaccharides Produced by Gluconobacteroxydans NCIMB 4943 by the Human Colonic Microflora. Curr Issues Intestinal Microbiol 7: 7-12. |
| [93] |
Gibson GR, Beatty ER, Wang X, et al. (1995) Selective stimulation of bifidobacteria in the human colon by oligofructose and inulin. Gastroenterology 108: 975-982. doi: 10.1016/0016-5085(95)90192-2
|
| [94] |
Bouhnik Y, Achour L, Paineau D, et al. (2007) Four-week short chain fructo-oligosaccharides ingestion leads to increasing fecal bifidobacteria and cholesterol excretion in healthy elderly volunteers. Nutrition 6: 42-46. doi: 10.1186/1475-2891-6-42
|
| [95] | Depeint F, Tzortzis G, Vulevic J, et al. (2008) Prebiotic evaluation of a novel galactooligosaccharide mixture produced by the enzymatic activity of Bifidobacterium bifidum NCIMB 41171, in healthy humans: a randomized, double-blind, crossover, placebo-controlled intervention study. Am J Clin Nutr 87: 785-791. |
| [96] | Kleessen B, Sykura B, Zunft HJ, et al. (1997) Effects of inulin and lactose on fecal microflora, microbial activity, and bowel habit in elderly constipated persons. Am J Clin Nutr 65: 1397-1402. |
| [97] | Kruse HP, Kleessen B, Blaut M (1999) Effects of inulin on faecal bifidobacteria in human subjects. Br J Nutr 82: 375-382. |
| [98] |
Ladirat SE, Schoterman MH, Rahaoui H, et al. (2014) Exploring the effects of galacto-oligosaccharides on the gut microbiota of healthy adults receiving amoxicillin treatment. Br J Nutr 112: 536-546. doi: 10.1017/S0007114514001135
|
| [99] |
Petry N, Egli I, Chassard C, et al. (2012) Inulin modifies the bifidobacteria population, fecal lactate concentration, and fecal pH but does not influence iron absorption in women with low iron status. Am J Clin Nutr 96: 325-331. doi: 10.3945/ajcn.112.035717
|
| [100] |
Tuohy KM, Kolida S, Lustenberger AM, et al. (2001) The prebiotic effects of biscuits containing partially hydrolysed guar gum and fructo-oligosaccharides - a human volunteer study. Br J Nutr 86: 341-348. doi: 10.1079/BJN2001394
|
| [101] |
Bäckhed F, Ding H, Wang T, et al. (2004) The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci U S A 101: 15718-15723. doi: 10.1073/pnas.0407076101
|
| [102] |
Tenbaum SP, Ordóñez-Morán P, Puig I, et al. (2012) β-catenin confers resistance to PI3K and AKT inhibitors and subverts FOXO3a to promote metastasis in colon cancer. Nat Med 18: 892-901. doi: 10.1038/nm.2772
|
| [103] | Leenen CH, Dieleman LA (2007) Inulin and oligofructose in chronic inflammatory bowel disease. J Nutr 137: 2572S-2575S. |
| [104] |
Puertollano E, Kolida S, Yaqoob P (2014) Biological significance of short-chain fatty acid metabolism by the intestinal microbiome. Curr Opin Clin Nutr Metab Care 17: 139-144. doi: 10.1097/MCO.0000000000000025
|
| [105] |
Canavan C, Abrams KR, Mayberry J (2006) Meta-analysis: colorectal and small bowel cancer risk in patients with Crohn's disease. Aliment Pharmacol Ther 23: 1097-1104. doi: 10.1111/j.1365-2036.2006.02854.x
|
| [106] |
Lindsay JO, Whelan K, Stagg AJ, et al. (2006) Clinical, microbiological and immunological effects of fructooligosaccharide in patients with Crohn's disease. Gut 55: 348-355. doi: 10.1136/gut.2005.074971
|
| [107] |
Candela M, Turroni S, Biagi E, et al. (2014) Inflammation and colorectal cancer, when microbiota-host mutualism breaks. World J Gastroenterol 20: 908-922. doi: 10.3748/wjg.v20.i4.908
|
| [108] |
Silk DB, Davis A, Vulevic J, et al. (2009) Clinical trial: the effects of a trans-galactooligosaccharide prebiotic on faecal microbiota and symptoms in irritable bowel syndrome. Aliment Pharmacol Ther 29: 508-518. doi: 10.1111/j.1365-2036.2008.03911.x
|
| [109] |
Whelan K (2011) Probiotics and prebiotics in the management of irritable bowel syndrome: a review of recent clinical trials and systematic reviews. Curr Opin Clin Nutr Metab Care 14: 581-587. doi: 10.1097/MCO.0b013e32834b8082
|
| [110] | Louis P, Hold GL, Flint HJ (2014) The gut microbiota, bacterial metabolites and colorectal cancer. Nat Rev Microbiol12: 661-672. |
| [111] |
Scanlan PD, Shanahan F, Clune Y, et al. (2008) Culture-independent analysis of the gut microbiota in colorectal cancer and polyposis. Environ Microbiol 10: 789-798. doi: 10.1111/j.1462-2920.2007.01503.x
|
| [112] |
Marchesi JR, Dutilh BE, Hall N, et al. (2011) Towards the human colorectal cancer microbiome. PLoS One 6: e20447. doi: 10.1371/journal.pone.0020447. doi: 10.1371/journal.pone.0020447
|
| [113] |
Shen XJ, Rawls JF, Randall T, et al. (2010) Molecular characterization of mucosal adherent bacteria and associations with colorectal adenomas. Gut Microbes 1: 138-147. doi: 10.4161/gmic.1.3.12360. doi: 10.4161/gmic.1.3.12360
|
| [114] |
Sobhani I, Tap J, Roudot-Thoraval F, et al. (2011) Microbial dysbiosis in colorectal cancer (CRC) patients. PLoS One 6: e16393. doi: 10.1371/journal.pone.0016393
|
| [115] | Moore WE, Moore LH (1995) Intestinal floras of populations that have a high risk of colorectal cancer. Appl Environ Microbiol 61: 3202-3207. |
| [116] |
Dewulf EM, Cani PD, Claus SP, et al. (2013) Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 62: 1112-1121. doi: 10.1136/gutjnl-2012-303304
|
| [117] | Bosscher D, Breynaert A, Pieters L, et al. (2009). Food-based strategies to modulate the composition of the intestinal microbiota and their associated health effects. J Physiol Pharmacol 60(Suppl 6):5-11. |
| [118] | Roberfroid MB (2007) Inulin-type fructans: functional food ingredients. J Nutr 137: 2493S-2502S. |
| [119] | Canfora EE, Jocken JW, Blaak EE (2015) Short-chain fatty acids in control of body weight and insulin sensitivity. Nat Rev Endocrinol 11: 5775-5791. |
| [120] | Hamer HM, Jonkers D, Venema K, et al. (2008). Review article: the role of butyrate on colonic function. Aliment Pharmacol Ther 27: 104-119. |
| [121] |
Wong JM, de Souza R, Kendall CW, et al. (2006) Colonic health: fermentation and short chain fatty acids. J Clin Gastroenterol 40: 235-243. doi: 10.1097/00004836-200603000-00015
|
| [122] |
Tilg H, Moschen AR (2014) Microbiota and diabetes: an evolving relationship. Gut 63: 1513-1521 doi: 10.1136/gutjnl-2014-306928
|
| [123] |
Wagner NM, Brandhorst G, Czepluch F, et al. (2013) Circulating regulatory T cells are reduced in obesity and may identify subjects at increased metabolic and cardiovascular risk. Obesity Silver Spring 21: 461-468. doi: 10.1002/oby.20087
|
| [124] | Duncan SH, Aminov RI, Scott KP, et al. (2006) Proposal of Roseburia faecis sp. nov., Roseburia hominis sp. nov., and Roseburia inulinivorans sp. nov., based on isolates from human faeces. Int J Syst Evol Micr 56: 2437-2441. |
| [125] | Duncan SH, Hold GL, Harmsen HJM, et al. (2002) Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen nov., comb. nov. Int J Syst Evol Micr 52: 2141-2146. |
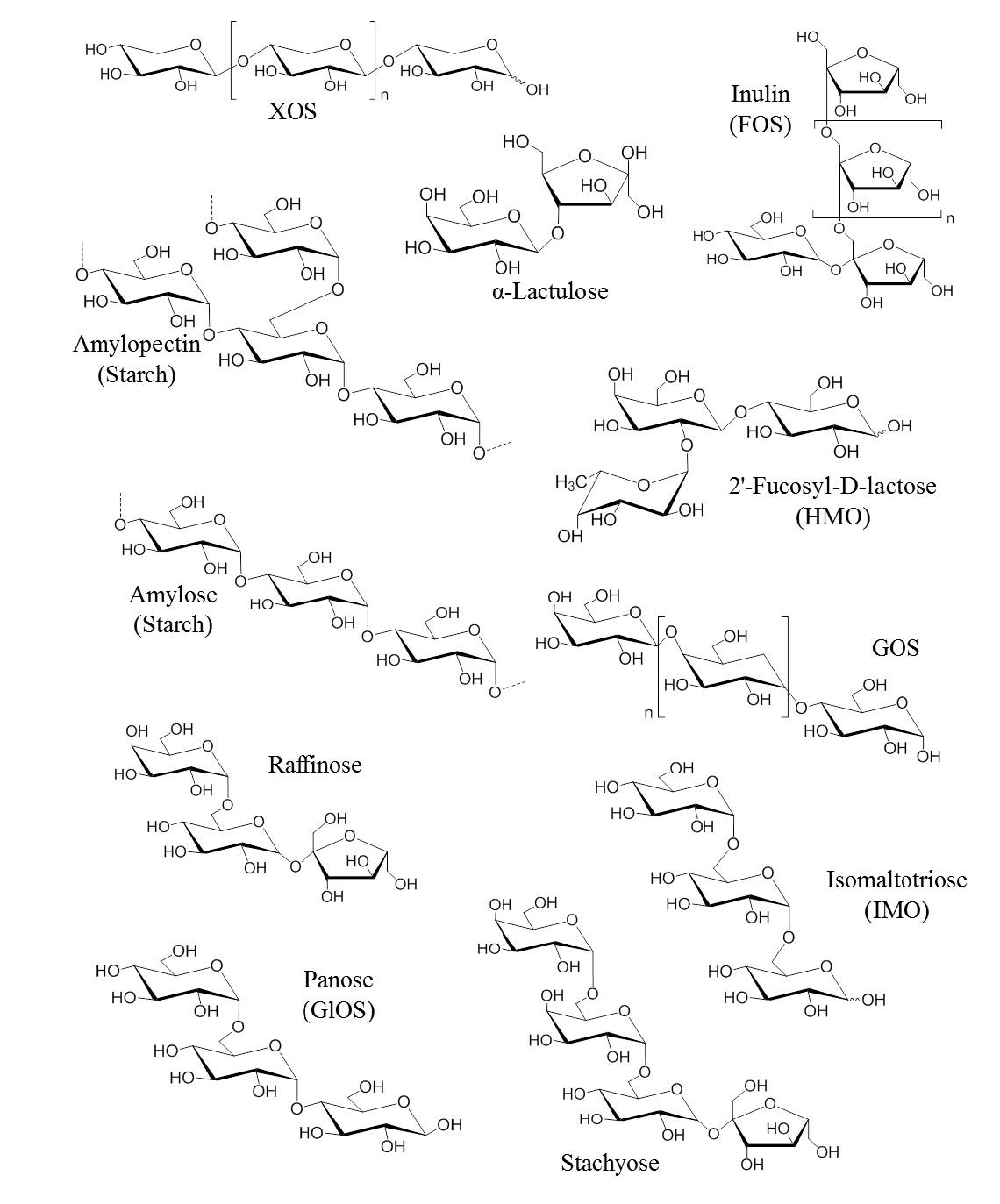
Figures(1)

Javier Fernández, Sául Redondo-Blanco, Elisa M. Miguélez, Claudio J. Villar, Alfonso Clemente, Felipe Lombó. Healthy effects of prebiotics and their metabolites against intestinal diseases and colorectal cancer[J]. AIMS Microbiology, 2015, 1(1): 48-71. doi: 10.3934/microbiol.2015.1.48







 DownLoad:
DownLoad: