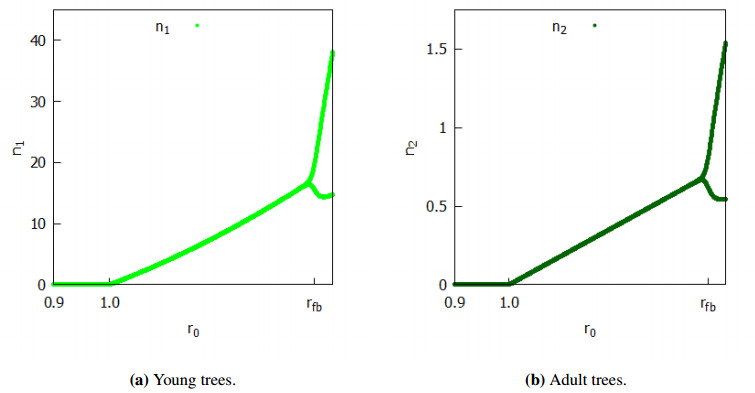
The work presented a general discrete-time model of a population of trees affected by a parasite. The tree population was considered size-structured, and the parasite was represented by a single scalar variable. Parasite dynamics were assumed to act on a faster timescale than tree dynamics. The model was studied based on an associated nonlinear matrix model, in which the presence of the parasites was only reflected in the value of its parameters. For the model in all its generality, an explicit condition of viability/extinction of the parasite/tree community was found. In a simplified model with two size-classes of trees and particular forms of the vital rates, it was shown that the model undergoes a transcritical bifurcation and, likewise, a period-doubling bifurcation. It was found that, for any tree fertility rate that makes them viable without a parasite, if the parasite sufficiently reduces the survival of young trees, it can lead to the extinction of the entire community. The same cannot be assured if the parasite acts on adult trees. In situations where a high fertility rate coupled with a low survival rate of adult trees causes a non-parasitized population of trees to fluctuate, a parasite sufficiently damaging only young trees can stabilize the population. If, instead, the parasite acts on adult trees, we can find a destabilization condition on the tree population that brings them from a stable to an oscillating regime.
Citation: Rafael Bravo de la Parra, Ezio Venturino. A discrete two time scales model of a size-structured population of parasitized trees[J]. Mathematical Biosciences and Engineering, 2024, 21(9): 7040-7066. doi: 10.3934/mbe.2024309
The work presented a general discrete-time model of a population of trees affected by a parasite. The tree population was considered size-structured, and the parasite was represented by a single scalar variable. Parasite dynamics were assumed to act on a faster timescale than tree dynamics. The model was studied based on an associated nonlinear matrix model, in which the presence of the parasites was only reflected in the value of its parameters. For the model in all its generality, an explicit condition of viability/extinction of the parasite/tree community was found. In a simplified model with two size-classes of trees and particular forms of the vital rates, it was shown that the model undergoes a transcritical bifurcation and, likewise, a period-doubling bifurcation. It was found that, for any tree fertility rate that makes them viable without a parasite, if the parasite sufficiently reduces the survival of young trees, it can lead to the extinction of the entire community. The same cannot be assured if the parasite acts on adult trees. In situations where a high fertility rate coupled with a low survival rate of adult trees causes a non-parasitized population of trees to fluctuate, a parasite sufficiently damaging only young trees can stabilize the population. If, instead, the parasite acts on adult trees, we can find a destabilization condition on the tree population that brings them from a stable to an oscillating regime.

| [1] | M. J. Hatcher, A. M. Dunn, Parasites in Ecological Communities. From Interactions to Ecosystems, Cambridge University Press, Cambridge, 2011. https://doi.org/10.1017/CBO9780511987359 |
| [2] |
D. Ludwig, D. D. Jones, C. S. Holling, Qualitative analysis of insect outbreak systems: the spruce budworm and forest, J. Anim. Ecol., 47 (1978), 315–332. https://doi.org/10.2307/3939 doi: 10.2307/3939

|
| [3] |
S. Rinaldi, S. Muratori, Limit cycles in slow-fast forest-pest models, Theor. Popul. Biol., 41 (1992), 26–43. https://doi.org/10.1016/0040-5809(92)90048-X doi: 10.1016/0040-5809(92)90048-X
|
| [4] |
P. Magal, Z. Zhang, Numerical simulations of a population dynamic model describing parasite destruction in a wild type pine forest, Ecol. Complex., 34 (2018), 147–160. https://doi.org/10.1016/j.ecocom.2017.05.001 doi: 10.1016/j.ecocom.2017.05.001
|
| [5] |
I. Tankam-Chedjou, S. Touzeau, L. Mailleret, J. J. Tewa, F. Grognard, Modelling and control of a banana soilborne pest in a multi-seasonal framework, Math. Biosci., 322 (2020), 108324. https://doi.org/10.1016/j.mbs.2020.108324 doi: 10.1016/j.mbs.2020.108324
|
| [6] |
T. M. M. De Silva, S. R. J. Jang, Period-doubling and Neimark-Sacker bifurcations in a larch budmoth population model, J. Differ. Equations Appl., 23 (2017), 1619–1639. https://doi.org/10.1080/10236198.2017.1354989. doi: 10.1080/10236198.2017.1354989
|
| [7] | M. Iannelli, A. Pugliese, An Introduction to Mathematical Population Dynamics. Along the trail of Volterra and Lotka, Springer, 2014. |
| [8] |
J. Liang, N. Picard, Matrix model of forest dynamics: An overview and outlook, Forest Sci., 59 (2013), 359–378. https://doi.org/10.5849/forsci.11-123 doi: 10.5849/forsci.11-123
|
| [9] |
F. Raffa, B. H. Aukema, B. J. Bentz, A. L. Carroll, J. A. Hicke, M. G. Turner, et al., Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions, BioScience, 58 (2008), 501–517. https://doi.org/10.1641/B580607 doi: 10.1641/B580607
|
| [10] |
L. Sanz, R. Bravo de la Parra, E. Sánchez, Approximate reduction of non-linear discrete models with two time scales, J. Differ. Equations Appl., 14 (2008), 607–627. https://doi.org/10.1186/s13662-019-2303-1 doi: 10.1186/s13662-019-2303-1
|
| [11] |
L. Sanz, R. Bravo de la Parra, M. Marvá, E. Sánchez, Non-linear population discrete models with two time scales: Re-scaling of part of the slow process, Adv. Differ. Equations, 2019 (2019), 401. https://doi.org/10.1186/s13662-019-2303-1 doi: 10.1186/s13662-019-2303-1
|
| [12] | J. M. Cushing, An Introduction to Structured Population Dynamics, CBMS-NSF Regional Conference Series in Applied Mathematics, Society for Industrial and Applied Mathematics, Philadelphia, 1998. https://doi.org/10.1137/1.9781611970005. |
| [13] | H. L. Smith, H. R. Thieme, Dynamical Systems and Population Persistence, American Mathematical Society, Providence, RI, 2011. https://doi.org/10.1090/gsm/118 |
| [14] | L. J. S. Allen, An Introduction to Mathematical Biology, Pearson Prentice Hall, Upper Saddle River, NJ, 2007. |
| [15] |
P. Cull, Population models: Stability in one dimension, Bull. Math. Biol., 69 (2007), 989–1017. https://doi.org/10.1007/s11538-006-9129-1 doi: 10.1007/s11538-006-9129-1
|
| [16] |
E. Liz, A new flexible discrete-time model for stable populations, Discrete Con. Dyn.-B, 23 (2018), 2487–2498. https://doi.org/10.3934/dcdsb.2018066 doi: 10.3934/dcdsb.2018066
|
| [17] |
M. A. Zavala, R. Bravo de la Parra, A mechanistic model of tree competition and facilitation for Mediterranean forests: Scaling from leaf physiology to stand dynamics, Ecol. Model., 188 (2005), 76–92. https://doi.org/10.1016/j.ecolmodel.2005.05.006 doi: 10.1016/j.ecolmodel.2005.05.006
|
| [18] |
R. Kon, Y. Saito, Y. Takeuchi, Permanence of single-species stage-structured models, J. Math. Biol., 48 (2004), 515–528. https://doi.org/10.1007/s00285-003-0239-1 doi: 10.1007/s00285-003-0239-1
|
| [19] |
R. N. Thompson, E. Brooks-Pollock, Detection, forecasting and control of infectious disease epidemics: modelling outbreaks in humans, animals and plants, Phil. Trans. R. Soc. B, 374 (2019), 20190038. https://doi.org/10.1098/rstb.2019.0038 doi: 10.1098/rstb.2019.0038
|
| [20] |
N. J. Cunniffe, B. Koskella, C. J. E. Metcalf, S. Parnell, T. R. Gottwald, C. A. Gilligan, Thirteen challenges in modelling plant diseases, Epidemics, 10 (2015), 6–10. https://doi.org/10.1016/j.epidem.2014.06.002 doi: 10.1016/j.epidem.2014.06.002
|
| [21] |
I. L. Boyd, P. H. Freer-Smith, C. A. Gilligan, H. C. J. Godfray, The consequence of tree pests and diseases for ecosystem services, Science, 342 (2013), 1235773. https://doi.org/10.1126/science.1235773 doi: 10.1126/science.1235773
|
| [22] |
R. N. Thompson, C. A. Gilligan, N. J. Cunniffe, Control fast or control smart: when should invading pathogens be controlled?, PLoS Comput. Biol., 14 (2018), 1–21. https://doi.org/10.1371/journal.pcbi.1006014 doi: 10.1371/journal.pcbi.1006014
|
| [23] | Y. A. Kuznetsov, Elements of Applied Bifurcation Theory, 3$^{rd}$ edition, Springer, New York, 2004. |


Figures(3) / Tables(1)

Rafael Bravo de la Parra, Ezio Venturino. A discrete two time scales model of a size-structured population of parasitized trees[J]. Mathematical Biosciences and Engineering, 2024, 21(9): 7040-7066. doi: 10.3934/mbe.2024309







 DownLoad:
DownLoad: