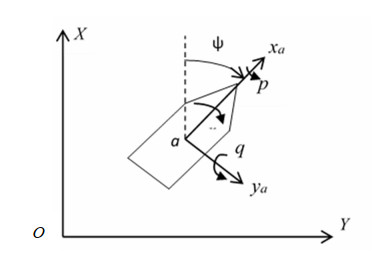
This article considered the sampled-data control issue for a dynamic positioning ship (DPS) with the Takagi-Sugeno (T-S) fuzzy model. By introducing new useful terms such as second-order term of time, an improved Lyapunov-Krasovskii function (LKF) was constructed. Additionally, the reciprocally convex method is introduced to bound the derivative of LKF. According to the constructed LKF, the sampling information during the whole sampling period was fully utilized, and less conservatism was obtained. Then, the stability condition, robust performance, mode uncertainty and sampled-data controller design were analyzed by means of the linear matrix inequality (LMI). Finally, an example was given to demonstrate the effectiveness of the proposed method.
Citation: Minjie Zheng, Yulai Su, Guoquan Chen. An improved sampled-data control for a nonlinear dynamic positioning ship with Takagi-Sugeno fuzzy model[J]. Mathematical Biosciences and Engineering, 2024, 21(5): 6019-6041. doi: 10.3934/mbe.2024265
This article considered the sampled-data control issue for a dynamic positioning ship (DPS) with the Takagi-Sugeno (T-S) fuzzy model. By introducing new useful terms such as second-order term of time, an improved Lyapunov-Krasovskii function (LKF) was constructed. Additionally, the reciprocally convex method is introduced to bound the derivative of LKF. According to the constructed LKF, the sampling information during the whole sampling period was fully utilized, and less conservatism was obtained. Then, the stability condition, robust performance, mode uncertainty and sampled-data controller design were analyzed by means of the linear matrix inequality (LMI). Finally, an example was given to demonstrate the effectiveness of the proposed method.

| [1] |
H. R. Karimi, Y. Lu, Guidance and control methodologies for marine vehicles: A survey, Control Eng. Pract., 111 (2021), 104785. https://doi.org/10.1016/j.conengprac.2021.104785 doi: 10.1016/j.conengprac.2021.104785

|
| [2] | T. I. Fossen, Handbook of Marine Craft Hydrodynamics and Motion Control, Wiley, 2011. |
| [3] | J. C. Patra, D. Wang, Ship dynamic positioning control system: A review, in Proceedings of the 2004 American Control Conference, (2004). |
| [4] |
K. D. Do, Global robust and adaptive output feedback control for a marine dynamic positioning of surface ships, J. Mar. Sci. Appl., 10 (2011), 325–332. https://doi.org/10.1007/s11804-011-1076-z doi: 10.1007/s11804-011-1076-z
|
| [5] |
Y. Su, C. Zheng, P. Mercorelli, Nonlinear PD fault-tolerant control for dynamic positioning of ships with actuator constraints, IEEE/ASME Trans. Mech., 22 (2017), 1132−31142. https://doi.org/10.1109/TMECH.2016.2603538 doi: 10.1109/TMECH.2016.2603538
|
| [6] |
G. Zhang, M. Yao, W. Zhang, W. Zhang, Event-triggered distributed adaptive cooperative control for multiple dynamic positioning ships with actuator faults, Ocean Eng., 242 (2021), 110124. https://doi.org/10.1016/j.oceaneng.2021.110124 doi: 10.1016/j.oceaneng.2021.110124
|
| [7] |
S. Donnarumma, M. Martelli, F. D'Agostino, D. Kaza, F. Silvestro, Multiphysics modeling and simulation of integrated electric propulsion system for ship dynamic positioning, IEEE Trans. Ind. Appl., 2024 (2024), 1–10. https://doi.org/10.1109/speedam53979.2022.9841976 doi: 10.1109/speedam53979.2022.9841976
|
| [8] |
Y. Wang, X. Yang, H. Yan, Reliable fuzzy tracking control of near-space hypersonic vehicle using aperiodic measurement information, IEEE Trans. Ind. Electron., 66 (2019), 9439–9447. https://doi.org/10.1109/TIE.2019.2892696 doi: 10.1109/TIE.2019.2892696
|
| [9] |
X. Meng, B. Jiang, H. R. Karimi, C. Gao, Leader-follower sliding mode formation control of fractional-order multi-agent systems: A dynamic event-triggered mechanism, Neurocomputing, 557 (2023), 126691. https://doi.org/10.1016/j.neucom.2023.126691 doi: 10.1016/j.neucom.2023.126691
|
| [10] |
Y. Wang, Y. Xia, P. Zhou, Fuzzy-model-based sampled-data control of chaotic systems: A fuzzy time-dependent Lyapunov-Krasovskii functional approach, IEEE Trans. Fuzzy Syst., 25 (2016), 1672–1684. https://doi.org/10.1109/TFUZZ.2016.2617378 doi: 10.1109/TFUZZ.2016.2617378
|
| [11] |
Y. Wang, P. Shi, On master-slave synchronization of Chaotic Lur'e systems using sampled-data control, IEEE Trans. Circuits Syst. II, 85 (2016), 981–992. https://doi.org/10.1007/s11071-016-2737-x doi: 10.1007/s11071-016-2737-x
|
| [12] |
W. H. Chen, Z. Wang, X. Lu, On sampled-data control for masterslave synchronization of chaotic Lur'e systems, IEEE Trans. Circuits Syst. II, 59 (2012), 515–519. https://doi.org/10.1109/TCSII.2012.2204114 doi: 10.1109/TCSII.2012.2204114
|
| [13] |
H. Xiao, Q. Zhu, H. R. Karimi, Stability of stochastic delay switched neural networks with all unstable subsystems: A multiple discretized Lyapunov-Krasovskii functionals method, Inf. Sci., 582 (2022), 302–315. https://doi.org/10.1016/j.ins.2021.09.027 doi: 10.1016/j.ins.2021.09.027
|
| [14] |
Z. G. Wu, P. Shi, H. Su, J. Chu, Stochastic synchronization of Markovian jump neural networks with time-varying delay using sampled data, IEEE Trans. Cybern., 43 (2013), 1796–1806. https://doi.org/10.1109/TSMCB.2012.2230441 doi: 10.1109/TSMCB.2012.2230441
|
| [15] |
Z. G. Wu, P. Shi, H. Su, J. Chu, Local synchronization of chaotic neural networks with sampled-data and saturating actuators, IEEE Trans. Cybern., 44 (2014), 2635–2645. https://doi.org/10.1109/TCYB.2014.2312004 doi: 10.1109/TCYB.2014.2312004
|
| [16] |
B. Jiang, H. R. Karimi, X. Zhang, Z. Wu, Adaptive neural-network-based sliding mode control of switching distributed delay systems with Markov jump parameters, Neural Networks, 165 (2023), 846–859. https://doi.org/10.1016/j.neunet.2023.06.022 doi: 10.1016/j.neunet.2023.06.022
|
| [17] |
F. Ding, T. Chen, Hierarchical identification of lifted state-space models for general dual-rate systems, IEEE Trans. Circuits Syst. I, 52 (2005), 1179–1187. https://doi.org/10.1109/TCSI.2005.849144 doi: 10.1109/TCSI.2005.849144
|
| [18] |
L. Hu, P. Shi, P. Frank, Robust sampled-data control for Markovian jump linear systems, Automatica, 42 (2006), 2025–2030. https://doi.org/10.1016/j.automatica.2006.05.029 doi: 10.1016/j.automatica.2006.05.029
|
| [19] |
Z. G. Wu, P. Shi, H. Y. Su, Stochastic synchronization of Markovian jump neural networks with time-varying delay using sampled data, IEEE Trans. Cybern., 43 (2013), 796–1806. https://doi.org/10.1109/TSMCB.2012.2230441 doi: 10.1109/TSMCB.2012.2230441
|
| [20] |
L. Yan, Z. Wang, M. Zhang, Y. Fan, Sampled-data control for mean-square exponential stabilization of memristive neural networks under deception attacks, Chaos Solitons Fractals, 174 (2023), 113787. https://doi.org/10.1016/j.chaos.2023.113787 doi: 10.1016/j.chaos.2023.113787
|
| [21] |
A. Yerudkar, E. Chatzaroulas, C. Del Vecchio, S. Moschoyiannis, Sampled-data control of probabilistic boolean control networks: A deep reinforcement learning approach, Inf. Sci., 619 (2023), 374–389. https://doi.org/10.1016/j.ins.2022.11.030 doi: 10.1016/j.ins.2022.11.030
|
| [22] |
S. Li, L. Yang, K. Li, Z. Gao, Robust sampled-data cruise control scheduling of high speed train, Transp. Res. Part C, 46 (2014), 274–283. https://doi.org/10.1016/j.trc.2014.06.004 doi: 10.1016/j.trc.2014.06.004
|
| [23] |
Y. Wang, Q. Wang, P. Zhou, D. Duan, Robust H∞ directional control for a sampled-data autonomous airship, J. Cent. South Univ., 21 (2014), 1339–1346. https://doi.org/10.1007/s11771-014-2071-8 doi: 10.1007/s11771-014-2071-8
|
| [24] |
M. Zheng, Y. Zhou, S. Yang, L. Li, Robust H∞ control of neutral system for sampled-data dynamic positioning ships, IMA J. Math. Control Inf., 36 (2019), 1325–1345. https://doi.org/10.1093/imamci/dny029 doi: 10.1093/imamci/dny029
|
| [25] |
Z. Zou, M. Zheng, Design and stabilization analysis of luxury cruise with dynamic positioning systems based on sampled-data control, Math. Biosci. Eng., 20 (2023), 14026–14045. https://doi.org/10.3934/mbe.2023626 doi: 10.3934/mbe.2023626
|
| [26] |
M. Zheng, Y. Su, S. Yang, L. Li, RReliable fuzzy dynamic positioning tracking controller for unmanned surface vehicles based on aperiodic measurement information, Int. J. Fuzzy Syst., 25 (2023), 358–368. https://doi.org/10.1007/s40815-022-01414-9 doi: 10.1007/s40815-022-01414-9
|
| [27] |
H. Zhang, D. Yang, T. Chai, Guaranteed cost networked control for T-S fuzzy systems with time delays, IEEE Trans. Syst. Man Cybern. Part C, 37 (2007), 160–172. https://doi.org/10.1109/tsmcc.2006.88698 doi: 10.1109/tsmcc.2006.88698
|
| [28] |
P. Mercorelli, Using fuzzy PD controllers for soft motions in a car-like robot, Adv. Sci. Technol. Eng. Syst. J., 3 (2018), 380–390. https://doi.org/10.25046/aj030646 doi: 10.25046/aj030646
|
| [29] |
R. Sakthivel, S. A. Karthick, B. Kaviarasan, F. Alzahrani, Dissipativity-based non-fragile sampled-data control design of interval type-2 fuzzy systems subject to random delays, ISA Trans., 83 (2018), 154–164. https://doi.org/10.1016/j.isatra.2018.08.017 doi: 10.1016/j.isatra.2018.08.017
|
| [30] |
Z. Du, Y. Kao, J. H. Park, New results for sampled-data control of interval type-2 fuzzy nonlinear systems, J. Franklin Inst., 357 (2020), 121–141. https://doi.org/10.1016/j.jfranklin.2019.09.035 doi: 10.1016/j.jfranklin.2019.09.035
|
| [31] |
G. Velmurugan, Y. H. Joo, Sampled-data control design for TS fuzzy system via quadratic function negative determination approach, IEEE Trans. Fuzzy Syst., 32 (2024), 979–988. https://doi.org/10.1109/tfuzz.2023.3316351 doi: 10.1109/tfuzz.2023.3316351
|
| [32] |
H. Li, Y. Liu, Y. Ma, Stability of TS fuzzy system under non-fragile sampled-data H∞ control using augmented Lyapunov-Krasovskii functional, J. Franklin Inst., 360 (2023), 3162–3188. https://doi.org/10.1016/j.jfranklin.2023.01.032 doi: 10.1016/j.jfranklin.2023.01.032
|
| [33] |
H. Katayama, Nonlinear sampled-data stabilization of dynamically positioned ships, IEEE Trans. Control Syst. Technol., 18 (2010), 463–468. https://doi.org/10.1109/TCST.2009.2014876 doi: 10.1109/TCST.2009.2014876
|
| [34] |
H. Katayama, H. Aoki, Straight-line trajectory tracking control for sampled-data underactuated ships, IEEE Trans. Control Syst. Technol., 22 (2014), 1638–1645. https://doi.org/10.1109/TCST.2013.2280717 doi: 10.1109/TCST.2013.2280717
|
| [35] |
M. Zheng, Y. Zhou, S. Yang, L. Li, Robust H∞ control of neutral system for sampled-data dynamic positioning ships, IMA J. Math. Control Inf., 36 (2019), 1325–1345. https://doi.org/10.1093/imamci/dny029 doi: 10.1093/imamci/dny029
|
| [36] | S. Yang, M. Zheng, H-infinity fault-tolerant control for dynamic positioning ships based on sampled-data, J. Control Eng. Appl. Inf., 20 (2018), 32–39. |
| [37] |
M. Zheng, Y. Zhou, S. Yang, Robust fuzzy sampled-data control for dynamic positioning ships, J. Shanghai Jiaotong Univ., 23 (2018), 209–217. https://doi.org/10.1007/s12204-018-1931-z doi: 10.1007/s12204-018-1931-z
|
| [38] | G. Chen, Y. Suo, M. Zheng, S. Yang, L. Li, Reliable tracking control of dynamic positioning ships based on aperiodic measurement information, J. Control Eng. Appl. Inf., 24 (2022), 80–89. |
| [39] |
P. G. Park, J. W. Ko, C. Jeong, Reciprocally convex approach to stability of systems with time-varying delays, Automatica, 47 (2011), 235–238. https://doi.org/10.1016/j.automatica.2010.10.014 doi: 10.1016/j.automatica.2010.10.014
|
| [40] |
F. Yang, H. Zhang, Y. Wang, An enhanced input-delay approach to sampled-data stabilization of T-S fuzzy systems via mixed convex combination, Nonlinear Dyn., 75 (2014), 501–512. https://doi.org/10.1007/s11071-013-1080-8 doi: 10.1007/s11071-013-1080-8
|
| [41] |
L. Xie, Output feedback H∞ control of systems with parameter uncertainty, Int. J. Control, 63 (1996), 741–750. https://doi.org/10.1080/00207179608921866 doi: 10.1080/00207179608921866
|
| [42] |
E. Fridman, A refined input delay approach to sampled-data control, Automatica, 46 (2010), 421–427. https://doi.org/10.1016/j.automatica.2009.11.017 doi: 10.1016/j.automatica.2009.11.017
|
| [43] |
J. Yoneyama, Robust sampled-data stabilization of uncertain fuzzy systems via input delay approach, Inf. Sci., 198 (2012), 169–176. https://doi.org/10.1016/j.ins.2012.02.007 doi: 10.1016/j.ins.2012.02.007
|
| [44] |
H. Zhang, D. Yang, T. Y. Chai, Guaranteed cost networked control for T-S fuzzy systems with time delays, IEEE Trans. Syst. Man Cybern. Part C, 37 (2007), 160–172. https://doi.org/10.1109/tsmcc.2006.886983 doi: 10.1109/tsmcc.2006.886983
|
| [45] |
H. Shao, Q. L. Han, Z. Zhang, X. Zhu, Sampling-interval-dependent stability for sampled-data systems with state quantization, Int. J. Robust Nonlinear Control, 24 (2014), 2995–3008. https://doi.org/10.1002/rnc.3038 doi: 10.1002/rnc.3038
|
| [46] |
E. Tannuri, A. Agostinho, H. Morishita, L. Moratelli, Dynamic positioning systems: An experimental analysis of sliding mode control, Control Eng. Pract., 18 (2010), 1121–1132. https://doi.org/10.1016/j.conengprac.2010.06.007 doi: 10.1016/j.conengprac.2010.06.007
|









Figures(10) / Tables(1)

Minjie Zheng, Yulai Su, Guoquan Chen. An improved sampled-data control for a nonlinear dynamic positioning ship with Takagi-Sugeno fuzzy model[J]. Mathematical Biosciences and Engineering, 2024, 21(5): 6019-6041. doi: 10.3934/mbe.2024265







 DownLoad:
DownLoad: