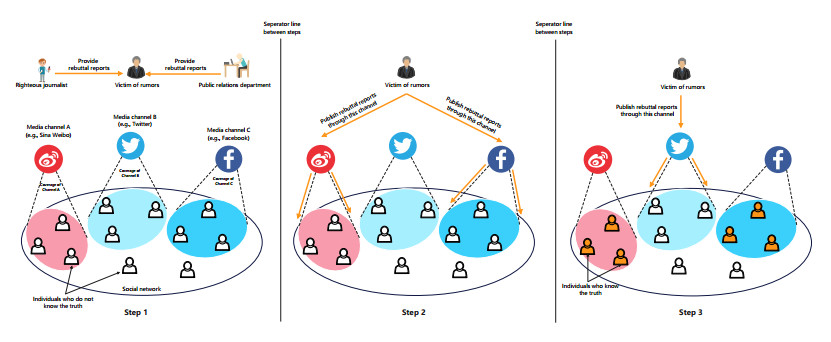
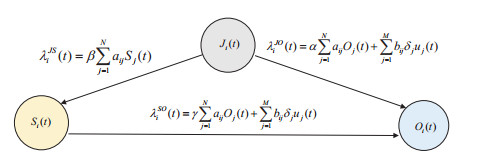
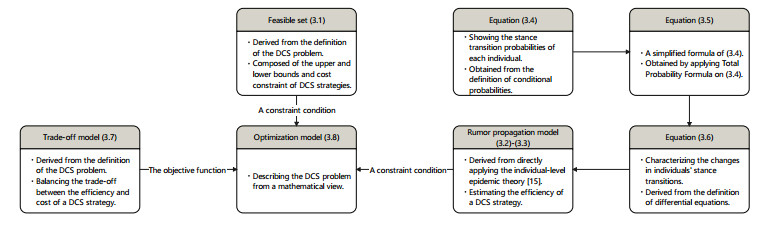
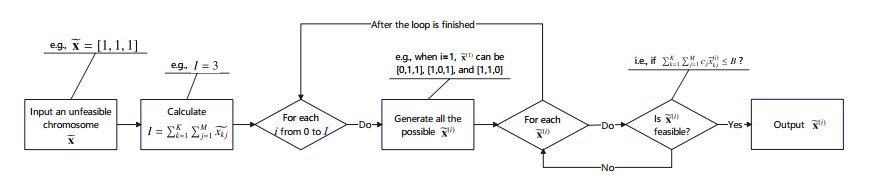
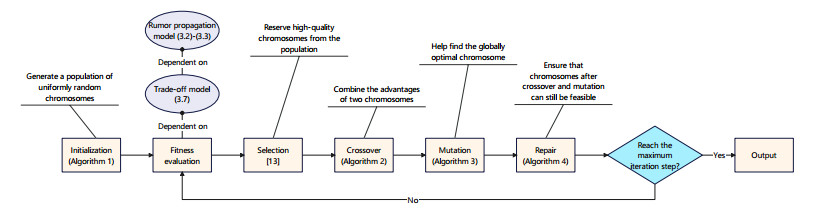
Rumors refer to spontaneously formed false stories. As rumors have shown severe threats to human society, it is significant to curb rumor propagation. Rumor clarification is an effective countermeasure on controlling rumor propagation. In this process, anti-rumor messages can be published through multiple media channels, including but not limited to online social platforms, TV programs and offline face-to-face campaigns. As the efficiency and cost of releasing anti-rumor information can vary from media channel to media channel, provided that the total budget is limited and fixed, it is valuable to investigate how to periodically select a combination of media channels to publish anti-rumor information so as to maximize the efficiency (i.e., make as many individuals as possible know the anti-rumor information) with the lowest cost. We refer to this issue as the dynamic channel selection (DCS) problem and any solution as a DCS strategy. To address the DCS problem, our contributions are as follows. First, we propose a rumor propagation model to characterize the influences of DCS strategies on curbing rumors. On this basis, we establish a trade-off model to evaluate DCS strategies and reduce the DCS problem to a mathematical optimization model called the DCS model. Second, based on the genetic algorithm framework, we develop a numerical method called the DCS algorithm to solve the DCS model. Third, we perform a series of numerical experiments to verify the performance of the DCS algorithm. Results show that the DCS algorithm can efficiently yield a satisfactory DCS strategy.
Citation: Yi Wang, Shicheng Zhong, Guo Wang. Dynamic selection of clarification channels in rumor propagation containment[J]. Mathematical Biosciences and Engineering, 2023, 20(8): 14995-15017. doi: 10.3934/mbe.2023672
Rumors refer to spontaneously formed false stories. As rumors have shown severe threats to human society, it is significant to curb rumor propagation. Rumor clarification is an effective countermeasure on controlling rumor propagation. In this process, anti-rumor messages can be published through multiple media channels, including but not limited to online social platforms, TV programs and offline face-to-face campaigns. As the efficiency and cost of releasing anti-rumor information can vary from media channel to media channel, provided that the total budget is limited and fixed, it is valuable to investigate how to periodically select a combination of media channels to publish anti-rumor information so as to maximize the efficiency (i.e., make as many individuals as possible know the anti-rumor information) with the lowest cost. We refer to this issue as the dynamic channel selection (DCS) problem and any solution as a DCS strategy. To address the DCS problem, our contributions are as follows. First, we propose a rumor propagation model to characterize the influences of DCS strategies on curbing rumors. On this basis, we establish a trade-off model to evaluate DCS strategies and reduce the DCS problem to a mathematical optimization model called the DCS model. Second, based on the genetic algorithm framework, we develop a numerical method called the DCS algorithm to solve the DCS model. Third, we perform a series of numerical experiments to verify the performance of the DCS algorithm. Results show that the DCS algorithm can efficiently yield a satisfactory DCS strategy.

| [1] |
X. Chen, N. Wang, Rumor spreading model considering rumor credibility, correlation and crowd classification based on personality, Sci. Rep., 10 (2020), 5887. https://doi.org/10.1038/s41598-020-62585-9 doi: 10.1038/s41598-020-62585-9

|
| [2] |
Z. Yu, S. Lu, D. Wang, Z. Li, Modeling and analysis of rumor propagation in social networks, Inform. Sci., 580 (2021), 857–873. https://doi.org/10.1016/j.ins.2021.09.012 doi: 10.1016/j.ins.2021.09.012
|
| [3] |
S. T. Malamut, M. Dawes, H. Xie, Characteristics of rumors and rumor victims in early adolescence: Rumor content and social impact, Soc. Dev., 27 (2018), 601–618. https://doi.org/10.1111/sode.12289 doi: 10.1111/sode.12289
|
| [4] |
P. Meel, D. K. Vishwakarma, Fake news, rumor, information pollution in social media and web: A contemporary survey of state-of-the-arts, challenges and opportunities, Exp. Syst. Appl., 153 (2020), 112986. https://doi.org/10.1016/j.eswa.2019.112986 doi: 10.1016/j.eswa.2019.112986
|
| [5] | T. Rana, P. Meel, P. Meel, Rumor propagation: A state-of-the-art survey of current challenges and opportunities, in 2019 2nd International Conference on Intelligent Communication and Computational Techniques (ICCT), (2019), 64–69. https://doi.org/10.1109/ICCT46177.2019.8969023 |
| [6] | J. A. Tucker, A. Guess, P. Barber, C. Vaccari, A. Siegel, S. Sanovich, et al., Social media, political polarization, and political disinformation: A review of the scientific literature, Political Polariz. Political Disinform., 2018. |
| [7] |
S. S. Ho, A. D. Leong, J. Looi, A. S. Chuah, Online, offline, or word-of-mouth? Complementary media usage patterns and credibility perceptions of nuclear energy information in Southeast Asia, Energy Res. Soc. Sci., 48 (2019), 46–56. https://doi.org/10.1016/j.erss.2018.09.012 doi: 10.1016/j.erss.2018.09.012
|
| [8] |
B. M. Okdie, R. E. Guadagno, F. J. Bernieri, A. L. Geers, A. R. Mclarney-Vesotski, Getting to know you: Face-to-face versus online interactions, Comput. Human Behav., 27 (2011), 153–159. https://doi.org/10.1016/j.chb.2010.07.017 doi: 10.1016/j.chb.2010.07.017
|
| [9] |
M. Paechter, B. Maier, Online or face-to-face? Students' experiences and preferences in e-learning, Int. Higher Educ., 13 (2010), 292–297. https://doi.org/10.1016/j.iheduc.2010.09.004 doi: 10.1016/j.iheduc.2010.09.004
|
| [10] |
M. Dohen, J. Schwartz, G. Bailly, Speech and face-to-face communication–An introduction, Speech Commun., 52 (2010), 477–480. https://doi.org/10.1016/j.specom.2010.02.016 doi: 10.1016/j.specom.2010.02.016
|
| [11] | M. Gollin, How much do social media ads cost on Facebook, Instagram, Twitter, and LinkedIn?, 2022. Available from: https://www.brandwatch.com/blog/how-much-do-social-media-ads-cost-on-facebook-instagram-twitter-and-linkedin/ |
| [12] | J. Kostka, Y. A. Oswald, R. Wattenhofer, Word of mouth: Rumor dissemination in social networks, in Structural Information and Communication Complexity: 15th International Colloquium, SIROCCO 2008 Villars-sur-Ollon, Springer, (2008), 185–196. https://doi.org/10.1007/978-3-540-69355-0_16 |
| [13] |
S. Katoch, S. S. Chauhan, V. Kumar, A review on genetic algorithm: past, present, and future, Multimedia Tools Appl., 80 (2021), 8091–8126. https://doi.org/10.1007/s11042-020-10139-6 doi: 10.1007/s11042-020-10139-6
|
| [14] |
D. P. Kroese, T. Brereton, T. Taimre, Z. I. Botev, Why the Monte Carlo method is so important today, Wiley Int. Rev. Comput. Stat., 6 (2014), 386–392. https://doi.org/10.1002/wics.1314 doi: 10.1002/wics.1314
|
| [15] |
V. Capasso, G. Serio, A generalization of the Kermack-McKendrick deterministic epidemic model, Math. Biosci., 42 (1978), 43–61. https://doi.org/10.1016/0025-5564(78)90006-8 doi: 10.1016/0025-5564(78)90006-8
|
| [16] |
P. Wan, X. Wang, X. Wang, L. Wang, Y. Lin, W. Zhao, Intervening coupling diffusion of competitive information in online social networks, IEEE Trans. Knowl. Data Eng., 33 (2019), 2548–2559. https://doi.org/10.1109/TKDE.2019.2954901 doi: 10.1109/TKDE.2019.2954901
|
| [17] |
X. Wang, X. Wang, F. Hao, G. Min, L. Wang, Efficient coupling diffusion of positive and negative information in online social networks, IEEE Trans. Network Serv. Manage., 16 (2019), 1226–1239. https://doi.org/10.1109/TNSM.2019.2917512 doi: 10.1109/TNSM.2019.2917512
|
| [18] |
F. Liu, M. Buss, Optimal control for heterogeneous node-based information epidemics over social networks, IEEE Trans. Control Network Syst., 7 (2020), 1115–1126. https://doi.org/10.1109/TCNS.2019.2963488 doi: 10.1109/TCNS.2019.2963488
|
| [19] |
L. Yang, T. Zhang, X. Yang, Y. Wu, Y. Y. Tang, Effectiveness analysis of a mixed rumor-quelling strategy, J. Franklin Inst., 355 (2018), 8079–8105. https://doi.org/10.1016/j.jfranklin.2018.07.040 doi: 10.1016/j.jfranklin.2018.07.040
|
| [20] |
Y. Xiao, W. Li, S. Qiang, Q. Li, H. Xiao, Y. Liu, A rumor & anti-rumor propagation model based on data enhancement and evolutionary game, IEEE Trans. Emerging Top. Comput., 10 (2020), 690–703. https://doi.org/10.1109/TETC.2020.3034188 doi: 10.1109/TETC.2020.3034188
|
| [21] |
Y. Xiao, Z. Huang, Q. Li, X. Lu, T. Li, Diffusion pixelation: A game diffusion model of rumor & anti-rumor inspired by image restoration, IEEE Trans. Knowl. Data Eng., 35 (2022), 4682–4694. https://doi.org/10.1109/TKDE.2022.3144310 doi: 10.1109/TKDE.2022.3144310
|
| [22] |
Q. Li, T. Xiang, T. Dai, Y. Xiao, An information dissemination model based on the rumor & anti-rumor & stimulate-rumor and tripartite cognitive game, IEEE Trans. Cognit. Dev. Syst., 15 (2022), 925–937. https://doi.org/10.1109/TCDS.2022.3193576 doi: 10.1109/TCDS.2022.3193576
|
| [23] |
C. Pan, L. Yang, X. Yang, Y. Wu, Y. Y. Tang, An effective rumor-containing strategy, Phys. A Stat. Mech. Appl., 500 (2018), 80–91. https://doi.org/10.1016/j.physa.2018.02.025 doi: 10.1016/j.physa.2018.02.025
|
| [24] |
P. Wan, X. Wang, X. Wang, L. Wang, Y. Lin, W. Zhao, Intervening coupling diffusion of competitive information in online social networks, IEEE Trans. Knowl. Data Eng., 33 (2019), 2548–2559. https://doi.org/10.1109/TKDE.2019.2954901 doi: 10.1109/TKDE.2019.2954901
|
| [25] |
Y. Lin, X. Wang, F. Hao, Y. Jiang, Y. Wu, G. Min, et al., Dynamic control of fraud information spreading in mobile social networks, IEEE Trans. Syst. Man, Cybern., 51 (2019), 3725–3738. https://doi.org/10.1109/TSMC.2019.2930908 doi: 10.1109/TSMC.2019.2930908
|
| [26] |
F. Liu, M. Buss, Optimal control for heterogeneous node-based information epidemics over social networks, IEEE Trans. Control Network Syst., 7 (2020), 1115–1126. https://doi.org/10.1109/TCNS.2019.2963488 doi: 10.1109/TCNS.2019.2963488
|
| [27] |
J. Chen, L. Yang, X. Yang, Y. Y. Tang, Cost-effective anti-rumor message-pushing schemes, Phys. A Stat. Mech. Appl., 540 (2020), 123085. https://doi.org/10.1016/j.physa.2019.123085 doi: 10.1016/j.physa.2019.123085
|
| [28] |
S. Wen, J. Jiang, Y. Xiang, S. Yu, W. Zhou, W. Jia, To shut them up or to clarify: Restraining the spread of rumors in online social networks, IEEE Trans. Parallel Distrib. Syst., 25 (2014), 3306–3316. https://doi.org/10.1109/TPDS.2013.2297115 doi: 10.1109/TPDS.2013.2297115
|
| [29] | M. Mohsin, Ten social media statistics you need to know in 2022, 2022. Available from: https://www.oberlo.com/blog/social-media-marketing-statistics. |
| [30] | N. Kin Wai, S. Horawalavithana, A. Iamnitchi, Multi-platform Information Operations: Twitter, Facebook and YouTube against the White Helmets, in Proceedings of The Workshop Proceedings of the 14th International AAAI Conference on Web and Social Media (ICWSM), 2021. |
| [31] |
D. A. Vega-Oliveros, L. da Fontoura Costa, F. A. Rodrigues, Influence maximization by rumor spreading on correlated networks through community identification, Commun. Nonlinear Sci. Numer. Simul., 83 (2020), 105094. https://doi.org/10.1016/j.cnsns.2019.105094 doi: 10.1016/j.cnsns.2019.105094
|
| [32] |
J. Chen, N. Wei, H. Yang, Immune Algorithm to Suppress Rumor Propagation Based on Influence Maximization, Secur. Commun. Networks, 2022 (2022). https://doi.org/10.1155/2022/6785828 doi: 10.1155/2022/6785828
|
| [33] |
K. Tanınmış, N. Aras, İ. Kuban Altınel, E. Güney, Minimizing the misinformation spread in social networks, Iise Trans., 52 (2020), 850–863. https://doi.org/10.1080/24725854.2019.1680909 doi: 10.1080/24725854.2019.1680909
|
| [34] |
L. Li, K. Huang, X. Yang, Containing Misinformation Spread: A Collaborative Resource Allocation Strategy for Knowledge Popularization and Expert Education, Secur. Commun. Networks, 2022 (2022), 1–14. https://doi.org/10.1155/2022/4510694 doi: 10.1155/2022/4510694
|
| [35] |
J. Zhu, P. Ni, G. Wang, Y. Li, Misinformation influence minimization problem based on group disbanded in social networks, Inform. Sci., 572 (2021), 1–15. https://doi.org/10.1016/j.ins.2021.04.086 doi: 10.1016/j.ins.2021.04.086
|
| [36] |
P. Ni, J. Zhu, G. Wang, Misinformation influence minimization by entity protection on multi-social networks, Appl. Intell., 0 (2022), 1–20. https://doi.org/10.1007/s10489-022-03798-5 doi: 10.1007/s10489-022-03798-5
|
| [37] |
L. Zino, M. Cao, Analysis, prediction, and control of epidemics: A survey from scalar to dynamic network models, IEEE Circuits Syst. Mag., 21 (2021), 4–23. https://doi.org/10.1109/MCAS.2021.3118100 doi: 10.1109/MCAS.2021.3118100
|
| [38] |
D. Huang, L. Yang, P. Li, X. Yang, Y. Y. Tang, Developing cost-effective rumor-refuting strategy through game-theoretic approach, IEEE Syst. J., 15 (2020), 5034–5045. https://doi.org/10.1109/JSYST.2020.3020078 doi: 10.1109/JSYST.2020.3020078
|
| [39] |
J. Zhao, L. Yang, X. Zhong, X. Yang, Y. Wu, Y. Y. Tang, Minimizing the impact of a rumor via isolation and conversion, Phys. A Stat. Mech. Appl., 526 (2019), 120867. https://doi.org/10.1016/j.physa.2019.04.103 doi: 10.1016/j.physa.2019.04.103
|
| [40] |
L. Yang, T. Zhang, X. Yang, Y. Wu, and Y. Y. Tang, Effectiveness analysis of a mixed rumor-quelling strategy, Journal of the Franklin Institute, 355 (2018), 8079–8105. https://doi.org/10.1016/j.jfranklin.2018.07.040 doi: 10.1016/j.jfranklin.2018.07.040
|
| [41] |
L. Yang, P. Li, X. Yang, Y. Xiang, Y. Y. Tang, Simultaneous benefit maximization of conflicting opinions: Modeling and analysis, IEEE Syst. J., 14 (2020), 1623–1634. https://doi.org/10.1109/JSYST.2020.2964004 doi: 10.1109/JSYST.2020.2964004
|
| [42] | C. Shyalika, An insight to genetic algorithms: Population initialization in genetic algorithms, 2019. Available from: https://medium.datadriveninvestor.com/population-initialization-in-genetic-algorithms-ddb037da6773 |
| [43] |
G. Marsaglia, A. Zaman, W. W. Tsang, Toward a universal random number generator, Stat. Probab. Lett., 9 (1990), 35–39. https://doi.org/10.1016/0167-7152(90)90092-L doi: 10.1016/0167-7152(90)90092-L
|
| [44] |
D. Bunnag, M. Sun, Genetic algorithm for constrained global optimization in continuous variables, Appl. Math. Comput., 171 (2005), 604–636. https://doi.org/10.1016/j.amc.2005.01.075 doi: 10.1016/j.amc.2005.01.075
|
| [45] |
A. Bodaghi, J. Oliveira, The characteristics of rumor spreaders on Twitter: A quantitative analysis on real data, Comput. Commun., 160 (2020), 674–687. https://doi.org/10.1016/j.comcom.2020.07.017 doi: 10.1016/j.comcom.2020.07.017
|
| [46] | A. Sonnenberg, How to Run Twitter Ads in 2022, 2022. Available from: https://www.socialmediaexaminer.com/how-to-run-twitter-ads-in-2022/. |
| [47] | S. Asur, B. A. Huberman, G. Szabo, C. Wang, Trends in social media: Persistence and decay, in Proceedings of the International AAAI Conference on Web and Social Media, (2011), 434–437. https://doi.org/10.2139/ssrn.1755748 |
Figures(7) / Tables(4)

Yi Wang, Shicheng Zhong, Guo Wang. Dynamic selection of clarification channels in rumor propagation containment[J]. Mathematical Biosciences and Engineering, 2023, 20(8): 14995-15017. doi: 10.3934/mbe.2023672







 DownLoad:
DownLoad: