Tumor hypoxia is commonly recognized as a condition stimulating the progress of the aggressive phenotype of tumor cells. Hypoxic tumor cells inhibit the delivery of cytotoxic drugs, causing hypoxic areas to receive insufficient amounts of anticancer agents, which results in adverse treatment responses. Being such an obstruction to conventional therapies for cancer, hypoxia might be considered a target to facilitate the efficacy of treatments in the resistive environment of tumor sites. In this regard, benefiting from prodrugs that selectively target hypoxic regions remains an effective approach. Additionally, combining hypoxia-activated prodrugs (HAPs) with conventional chemotherapeutic drugs has been used as a promising strategy to eradicate hypoxic cells. However, determining the appropriate sequencing and scheduling of the combination therapy is also of great importance in obtaining favorable results in anticancer therapy. Here, benefiting from a modeling approach, we study the efficacy of HAPs in combination with chemotherapeutic drugs on tumor growth and the treatment response. Different treatment schedules have been investigated to see the importance of determining the optimal schedule in combination therapy. The effectiveness of HAPs in varying hypoxic conditions has also been explored in the study. The model provides qualitative conclusions about the treatment response, as the maximal benefit is obtained from combination therapy with greater cell death for highly hypoxic tumors. It has also been observed that the antitumor effects of HAPs show a hypoxia-dependent profile.
Citation: Defne Yilmaz, Mert Tuzer, Mehmet Burcin Unlu. Assessing the therapeutic response of tumors to hypoxia-targeted prodrugs with an in silico approach[J]. Mathematical Biosciences and Engineering, 2022, 19(11): 10941-10962. doi: 10.3934/mbe.2022511
Tumor hypoxia is commonly recognized as a condition stimulating the progress of the aggressive phenotype of tumor cells. Hypoxic tumor cells inhibit the delivery of cytotoxic drugs, causing hypoxic areas to receive insufficient amounts of anticancer agents, which results in adverse treatment responses. Being such an obstruction to conventional therapies for cancer, hypoxia might be considered a target to facilitate the efficacy of treatments in the resistive environment of tumor sites. In this regard, benefiting from prodrugs that selectively target hypoxic regions remains an effective approach. Additionally, combining hypoxia-activated prodrugs (HAPs) with conventional chemotherapeutic drugs has been used as a promising strategy to eradicate hypoxic cells. However, determining the appropriate sequencing and scheduling of the combination therapy is also of great importance in obtaining favorable results in anticancer therapy. Here, benefiting from a modeling approach, we study the efficacy of HAPs in combination with chemotherapeutic drugs on tumor growth and the treatment response. Different treatment schedules have been investigated to see the importance of determining the optimal schedule in combination therapy. The effectiveness of HAPs in varying hypoxic conditions has also been explored in the study. The model provides qualitative conclusions about the treatment response, as the maximal benefit is obtained from combination therapy with greater cell death for highly hypoxic tumors. It has also been observed that the antitumor effects of HAPs show a hypoxia-dependent profile.

| [1] |
P. Vaupel, A. Mayer, Hypoxia in cancer: significance and impact on clinical outcome, Cancer Metastasis Rev., 26 (2007), 225–239. https://doi.org/10.1007/s10555-007-9055-1 doi: 10.1007/s10555-007-9055-1

|
| [2] |
C. T. Lee, M. K. Boss, M. W. Dewhirst, Imaging tumor hypoxia to advance radiation oncology, Antioxid. Redox Signaling, 21 (2014), 313–337. https://doi.org/10.1089/ars.2013.5759 doi: 10.1089/ars.2013.5759
|
| [3] |
M. R. Horsman, J. Overgaard, The impact of hypoxia and its modification of the outcome of radiotherapy, J. Radiat. Res., 57 (2016), i90–i98. https://doi.org/10.1093/jrr/rrw007 doi: 10.1093/jrr/rrw007
|
| [4] |
J. A. Forsythe, B. H. Jiang, N. V. Iyer, F. Agani, S. W. Leung, R. D. Koos, et al., Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1, Mol. Cell. Biol., 16 (1996), 4604–4613. https://doi.org/10.1128/MCB.16.9.4604 doi: 10.1128/MCB.16.9.4604
|
| [5] |
A. Ahluwalia, A. S Tarnawski, Critical role of hypoxia sensor-hif-1$\alpha$ in vegf gene activation. implications for angiogenesis and tissue injury healing, Curr. Med. Chem., 19 (2012), 90–97. https://doi.org/10.2174/092986712803413944 doi: 10.2174/092986712803413944
|
| [6] |
E. K. Rofstad, Microenvironment-induced cancer metastasis, Int. J. Radiat. Biol., 76 (2000), 589–605. https://doi.org/10.1080/095530000138259 doi: 10.1080/095530000138259
|
| [7] |
M. G. Binker, A. A. Binker-Cosen, D. Richards, H. Y. Gaisano, R. H. de Cosen, L. I. Cosen-Binker, Hypoxia–reoxygenation increase invasiveness of panc-1 cells through rac1/mmp-2, Biochem. Biophys. Res. Commun., 393 (2010), 371–376. https://doi.org/10.1016/j.bbrc.2010.01.125 doi: 10.1016/j.bbrc.2010.01.125
|
| [8] |
R. M. Phillips, Targeting the hypoxic fraction of tumours using hypoxia-activated prodrugs, Cancer Chemother. Pharmacol., 77 (2016), 441–457. https://doi.org/10.1007/s00280-015-2920-7 doi: 10.1007/s00280-015-2920-7
|
| [9] |
W. A. Denny, The role of hypoxia-activated prodrugs in cancer therapy, Lancet Oncol., 1 (2000), 25–29. https://doi.org/10.1016/S1470-2045(00)00006-1 doi: 10.1016/S1470-2045(00)00006-1
|
| [10] |
S. G. Peeters, C. M. Zegers, R. Biemans, N. G. Lieuwes, R. G. van Stiphout, A. Yaromina, et al., TH-302 in combination with radiotherapy enhances the therapeutic outcome and is associated with pretreatment [18f] hx4 hypoxia pet imaging, Clin. Cancer Res., 21 (2015), 2984–2992. https://doi.org/10.1158/1078-0432.CCR-15-0018 doi: 10.1158/1078-0432.CCR-15-0018
|
| [11] |
V. Liapis, A. Labrinidis, I. Zinonos, S. Hay, V. Ponomarev, V. Panagopoulos, et al., Hypoxia-activated pro-drug th-302 exhibits potent tumor suppressive activity and cooperates with chemotherapy against osteosarcoma, Cancer Lett., 357 (2015), 160–169. https://doi.org/10.1016/j.canlet.2014.11.020 doi: 10.1016/j.canlet.2014.11.020
|
| [12] |
Q. Liu, J. D. Sun, J. Wang, D. Ahluwalia, A. F. Baker, L. D. Cranmer, et al., TH-302, a hypoxia-activated prodrug with broad in vivo preclinical combination therapy efficacy: optimization of dosing regimens and schedules, Cancer Chemother. Pharmacol., 69 (2012), 1487–1498. https://doi.org/10.1007/s00280-012-1852-8 doi: 10.1007/s00280-012-1852-8
|
| [13] | Hypoxia/Normoxia, in Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine, Springer, Berlin, Heidelberg, (2006), 853–853. https://doi.org/10.1007/3-540-29623-9_7440 |
| [14] |
C. Wigerup, S. Påhlman, D. Bexell, Therapeutic targeting of hypoxia and hypoxia-inducible factors in cancer, Pharmacol. Ther., 164 (2016), 152–169. https://doi.org/10.1016/j.pharmthera.2016.04.009 doi: 10.1016/j.pharmthera.2016.04.009
|
| [15] |
J. D. Sun, Q. Liu, J. Wang, D. Ahluwalia, D. Ferraro, Y. Wang, et al., Selective tumor hypoxia targeting by hypoxia-activated prodrug TH-302 inhibits tumor growth in preclinical models of cancer, Clin. Cancer Res., 18 (2012), 758–770. https://doi.org/10.1158/1078-0432.CCR-11-1980 doi: 10.1158/1078-0432.CCR-11-1980
|
| [16] |
F. Meng, J. W. Evans, D. Bhupathi, M. Banica, L. Lan, G. Lorente, et al., Molecular and cellular pharmacology of the hypoxia-activated prodrug TH-302, Mol. Cancer Ther., 11 (2012), 740–751. https://doi.org/10.1158/1535-7163.MCT-11-0634 doi: 10.1158/1535-7163.MCT-11-0634
|
| [17] |
Y. Huang, Y. Tian, Y. Zhao, C. Xue, J. Zhan, L. Liu, et al., Efficacy of the hypoxia-activated prodrug evofosfamide (TH-302) in nasopharyngeal carcinoma in vitro and in vivo, Cancer Commun., 38 (2018), 1–9. https://doi.org/10.1186/s40880-018-0285-0 doi: 10.1186/s40880-018-0285-0
|
| [18] |
G. J. Weiss, J. R. Infante, E. G. Chiorean, M. J. Borad, J. C. Bendell, J. R. Molina, et al., Phase 1 study of the safety, tolerability, and pharmacokinetics of TH-302, a hypoxia-activated prodrug, in patients with advanced solid malignancies, Clin. Cancer Res., 17 (2011), 2997–3004. https://doi.org/10.1158/1078-0432.CCR-10-3425 doi: 10.1158/1078-0432.CCR-10-3425
|
| [19] |
I. Lohse, J. Rasowski, P. Cao, M. Pintilie, T. Do, M. S. Tsao, et al., Targeting hypoxic microenvironment of pancreatic xenografts with the hypoxia-activated prodrug th-302, Oncotarget, 7 (2016), 33571. https://doi.org/10.18632/oncotarget.9654 doi: 10.18632/oncotarget.9654
|
| [20] |
S. Matsumoto, S. Kishimoto, K. Saito, Y. Takakusagi, J. P. Munasinghe, N. Devasahayam, et al., Metabolic and physiologic imaging biomarkers of the tumor microenvironment predict treatment outcome with radiation or a hypoxia-activated prodrug in mice, Cancer Res., 78 (2018), 3783–3792. https://doi.org/10.1158/0008-5472.CAN-18-0491 doi: 10.1158/0008-5472.CAN-18-0491
|
| [21] |
K. J. Nytko, I. Grgic, S. Bender, J. Ott, M. Guckenberger, O. Riesterer et al., The hypoxia-activated prodrug evofosfamide in combination with multiple regimens of radiotherapy, Oncotarget, 8 (2017), 23702. https://doi.org/10.18632/oncotarget.15784 doi: 10.18632/oncotarget.15784
|
| [22] |
J. K. Saggar, I. F. Tannock, Activity of the hypoxia-activated pro-drug TH-302 in hypoxic and perivascular regions of solid tumors and its potential to enhance therapeutic effects of chemotherapy, Int. J. Cancer, 134 (2014), 2726–2734. https://doi.org/10.1002/ijc.28595 doi: 10.1002/ijc.28595
|
| [23] |
V. Liapis, I. Zinonos, A. Labrinidis, S. Hay, V. Ponomarev, V. Panagopoulos, et al., Anticancer efficacy of the hypoxia-activated prodrug evofosfamide (th-302) in osteolytic breast cancer murine models, Cancer Med., 5 (2016), 534–545. https://doi.org/10.1002/cam4.599 doi: 10.1002/cam4.599
|
| [24] |
J. K. Saggar, I. F. Tannock, Chemotherapy rescues hypoxic tumor cells and induces their reoxygenation and repopulation–-an effect that is inhibited by the hypoxia-activated prodrug TH-302, Clin. Cancer Res., 21 (2015), 2107–2114. https://doi.org/10.1158/1078-0432.CCR-14-2298 doi: 10.1158/1078-0432.CCR-14-2298
|
| [25] |
J. D. Sun, Q. Liu, D. Ahluwalia, W. Li, F. Meng, Y. Wang, et al., Efficacy and safety of the hypoxia-activated prodrug TH-302 in combination with gemcitabine and nab-paclitaxel in human tumor xenograft models of pancreatic cancer, Cancer Biol. Ther., 16 (2015), 438–449. https://doi.org/10.1080/15384047.2014.1003005 doi: 10.1080/15384047.2014.1003005
|
| [26] |
S. P. Chawla, L. D. Cranmer, B. A. Van Tine, D. R. Reed, S. H. Okuno, J. E. Butrynski, et al., Phase ii study of the safety and antitumor activity of the hypoxia-activated prodrug TH-302 in combination with doxorubicin in patients with advanced soft tissue sarcoma, J. Clin. Oncol., 32 (2014), 3299. https://doi.org/10.1200/JCO.2013.54.3660 doi: 10.1200/JCO.2013.54.3660
|
| [27] |
K. N. Ganjoo, L. D. Cranmer, J. E. Butrynski, D. Rushing, D. Adkins, S. H. Okuno, et al., A phase i study of the safety and pharmacokinetics of the hypoxia-activated prodrug th-302 in combination with doxorubicin in patients with advanced soft tissue sarcoma, Oncology, 80 (2011), 50–56. https://doi.org/10.1159/000327739 doi: 10.1159/000327739
|
| [28] |
J. Von Pawel, R. von Roemeling, U. Gatzemeier, M. Boyer, L. O. Elisson, P. Clark, et al., Tirapazamine plus cisplatin versus cisplatin in advanced non-small-cell lung cancer: A report of the international catapult I study group, J. Clin. Oncol., 18 (2000), 1351–1359. https://doi.org/10.1200/JCO.2000.18.6.1351 doi: 10.1200/JCO.2000.18.6.1351
|
| [29] |
F. W. Hunter, B. G. Wouters, W. R. Wilson, Hypoxia-activated prodrugs: paths forward in the era of personalised medicine, Br. J. Cancer, 114 (2016), 1071–1077. https://doi.org/10.1038/bjc.2016.79 doi: 10.1038/bjc.2016.79
|
| [30] | J. Sun, Q. Liu, D. Ahluwalia, J. Curd, M. Matteucci, C. Hart, Complementary chemotherapies with th-302, a novel hypoxia activated prodrug: optimization of dosing regimens and schedules for study in phase 1/2 with docetaxel, gemcitabine, pemetrexed, and doxorubicin, Biosymposia: Hypoxia, Ischemia, Inflammation, 2008. |
| [31] |
S. B. Reddy, S. K. Williamson, Tirapazamine: a novel agent targeting hypoxic tumor cells, Expert Opin. Invest. Drugs, 18 (2009), 77–87. https://doi.org/10.1517/13543780802567250 doi: 10.1517/13543780802567250
|
| [32] |
J. C. Forster, L. G. Marcu, E. Bezak, Approaches to combat hypoxia in cancer therapy and the potential for in silico models in their evaluation, Physica Med., 64 (2019), 145–156. https://doi.org/10.1016/j.ejmp.2019.07.006 doi: 10.1016/j.ejmp.2019.07.006
|
| [33] |
W. Tuckwell, E. Bezak, E. Yeoh, L. Marcu, Efficient Monte Carlo modelling of individual tumour cell propagation for hypoxic head and neck cancer, Phys. Med. Biol., 53 (2008), 4489. https://doi.org/10.1088/0031-9155/53/17/002 doi: 10.1088/0031-9155/53/17/002
|
| [34] |
W. M. Harriss-Phillips, E. Bezak, E. Yeoh, Monte Carlo radiotherapy simulations of accelerated repopulation and reoxygenation for hypoxic head and neck cancer, Br. J. Radiol., 84 (2011), 903–918. https://doi.org/10.1259/bjr/25012212 doi: 10.1259/bjr/25012212
|
| [35] |
W. M. Harriss-Phillips, E. Bezak, A. Potter, Stochastic predictions of cell kill during stereotactic ablative radiation therapy: Do hypoxia and reoxygenation really matter?, Int. J. Radiat. Oncol. Biol. Phys., 95 (2016), 1290–1297. https://doi.org/10.1016/j.ijrobp.2016.03.014 doi: 10.1016/j.ijrobp.2016.03.014
|
| [36] | L. G. Marcu, D. Marcu, S. M. Filip, In silico study of the impact of cancer stem cell dynamics and radiobiological hypoxia on tumour response to hyperfractionated radiotherapy, Cell proliferation, 49 (2016), 304–314. |
| [37] | E. Lindblom, I. Toma-Dasu, A. Dasu, Accounting for two forms of hypoxia for predicting tumour control probability in radiotherapy: An in silico study, in Oxygen Transport to Tissue XL. Advances in Experimental Medicine and Biology, 1072 (2018), 183–187. https://doi.org/10.1007/978-3-319-91287-5_29 |
| [38] |
A. Foehrenbacher, K. Patel, M. Abbattista, C. P. Guise, T. W. Secomb, W. R. Wilson, et al., The role of bystander effects in the antitumor activity of the hypoxia-activated prodrug pr-104, Frontiers in oncology, 3 (2013), 263. https://doi.org/10.3389/fonc.2013.00263 doi: 10.3389/fonc.2013.00263
|
| [39] |
A. B. Foehrenbacher, T. W. Secomb, W. R. Wilson, K. O. Hicks, Design of optimized hypoxia-activated prodrugs using pharmacokinetic/pharmacodynamic modeling, Front. Oncol., 3 (2013), 314. https://doi.org/10.3389/fonc.2013.00314 doi: 10.3389/fonc.2013.00314
|
| [40] |
K. O. Hicks, F. B. Pruijn, T. W. Secomb, M. P. Hay, R. Hsu, J. M. Brown, et al., Use of three-dimensional tissue cultures to model extravascular transport and predict in vivo activity of hypoxia-targeted anticancer drugs, J. Nat. Cancer Inst., 98 (2006), 1118–1128. https://doi.org/10.1093/jnci/djj306 doi: 10.1093/jnci/djj306
|
| [41] |
C. Meaney, G. G. Powathil, A. Yaromina, L. J. Dubois, P. Lambin, M. Kohandel, Role of hypoxia-activated prodrugs in combination with radiation therapy: An in silico approach, Math. Biosci. Eng., 16 (2019), 6257. https://doi.org/10.3934/mbe.2019312 doi: 10.3934/mbe.2019312
|
| [42] |
C. Meaney, S. Rhebergen, M. Kohandel, In silico analysis of hypoxia activated prodrugs in combination with anti angiogenic therapy through nanocell delivery, PLoS Comput. Biol., 16 (2020), e1007926. https://doi.org/10.1371/journal.pcbi.1007926 doi: 10.1371/journal.pcbi.1007926
|
| [43] |
S. Hamis, M. Kohandel, L. J. Dubois, A. Yaromina, P. Lambin, G. G. Powathil, Combining hypoxia-activated prodrugs and radiotherapy in silico: Impact of treatment scheduling and the intra-tumoural oxygen landscape, PLoS Comput. Biol., 16 (2020), e1008041. https://doi.org/10.1371/journal.pcbi.1008041 doi: 10.1371/journal.pcbi.1008041
|
| [44] |
X. Mao, S. McManaway, J. K. Jaiswal, P. B. Patel, W. R. Wilson, K. O. Hicks, et al., An agent-based model for drug-radiation interactions in the tumour microenvironment: Hypoxia-activated prodrug sn30000 in multicellular tumour spheroids, PLoS Comput. Biol., 14 (2018), e1006469. https://doi.org/10.1371/journal.pcbi.1006469 doi: 10.1371/journal.pcbi.1006469
|
| [45] |
C. R. Hong, S. Y. Mehta, H. Liyanage, S. P. McManaway, H. H. Lee, J. K. Jaiswal, et al., Spatially-resolved pharmacokinetic/pharmacodynamic modelling of bystander effects of a nitrochloromethylbenzindoline hypoxia-activated prodrug, Cancer Chemother. Pharmacol., 88 (2021), 673–687. https://doi.org/10.1007/s00280-021-04320-3 doi: 10.1007/s00280-021-04320-3
|
| [46] |
K. O. Hicks, B. G. Siim, J. K. Jaiswal, F. B. Pruijn, A. M. Fraser, R. Patel, et al., Pharmacokinetic/pharmacodynamic modeling identifies sn30000 and sn29751 as tirapazamine analogues with improved tissue penetration and hypoxic cell killing in tumors, Clin. Cancer Res., 16 (2010), 4946–4957. https://doi.org/10.1158/1078-0432.CCR-10-1439 doi: 10.1158/1078-0432.CCR-10-1439
|
| [47] |
C. R. Hong, W. R. Wilson, K. O. Hicks, An intratumor pharmacokinetic/pharmacodynamic model for the hypoxia-activated prodrug evofosfamide (TH-302): Monotherapy activity is not dependent on a bystander effect, Neoplasia, 21 (2019), 159–171. https://doi.org/10.1016/j.neo.2018.11.009 doi: 10.1016/j.neo.2018.11.009
|
| [48] |
C. R. Hong, G. Bogle, J. Wang, K. Patel, F. B. Pruijn, W. R. Wilson, et al., Bystander effects of hypoxia-activated prodrugs: agent-based modeling using three dimensional cell cultures, Front. Pharmacol., 9 (2018), 1013. https://doi.org/10.3389/fphar.2018.01013 doi: 10.3389/fphar.2018.01013
|
| [49] |
D. Lindsay, C. M. Garvey, S. M. Mumenthaler, J. Foo, Leveraging hypoxia-activated prodrugs to prevent drug resistance in solid tumors, PLoS Comput. Biol., 12 (2016), e1005077. https://doi.org/10.1371/journal.pcbi.1005077 doi: 10.1371/journal.pcbi.1005077
|
| [50] |
S. Yonucu, D. Yilmaz, C. Phipps, M. B. Unlu, M. Kohandel, Quantifying the effects of antiangiogenic and chemotherapy drug combinations on drug delivery and treatment efficacy, PLoS Comput. Biol., 13 (2017), 1–17, https://doi.org/10.1371/journal.pcbi.1005724 doi: 10.1371/journal.pcbi.1005724
|
| [51] |
M. Kohandel, M. Kardar, M. Milosevic, S. Sivaloganathan, Dynamics of tumor growth and combination of anti-angiogenic and cytotoxic therapies, Phys. Med. Biol., 52 (2007), 3665. https://doi.org/10.1088/0031-9155/52/13/001 doi: 10.1088/0031-9155/52/13/001
|
| [52] |
G. Powathil, M. Kohandel, S. Sivaloganathan, A. Oza, M. Milosevic, Mathematical modeling of brain tumors: effects of radiotherapy and chemotherapy, Phys. Med. Biol., 52 (2007), 3291. https://doi.org/10.1088/0031-9155/52/11/023 doi: 10.1088/0031-9155/52/11/023
|
| [53] |
E. Dalah, D. Bradley, A. Nisbet, A mathematical approach towards simulating a realistic tissue activity curve of 64Cu-ATSM for the purpose of sub-target volume delineation in radiotherapy, Nucl. Instrum. Methods Phys. Res., Sect. A, 619 (2010), 283–286. https://doi.org/10.1016/j.nima.2009.10.160 doi: 10.1016/j.nima.2009.10.160
|
| [54] | E. Dalah, D. Bradley, A. Nisbet, Simulation of tissue activity curves of 64Cu-ATSM for sub-target volume delineation in radiotherapy, Phys. Med. Biol., 55 (2010), 681. |
| [55] |
F. Winkler, S. V. Kozin, R. T. Tong, S. S. Chae, M. F. Booth, I. Garkavtsev, et al., Kinetics of vascular normalization by vegfr2 blockade governs brain tumor response to radiation: role of oxygenation, angiopoietin-1, and matrix metalloproteinases, Cancer Cell, 6 (2004), 553–563. https://doi.org/10.1016/j.ccr.2004.10.011 doi: 10.1016/j.ccr.2004.10.011
|
| [56] |
W. R. Wilson, M. P. Hay, Targeting hypoxia in cancer therapy, Nat. Rev. Cancer, 11 (2011), 393–410. https://doi.org/10.1038/nrc3064 doi: 10.1038/nrc3064
|
| [57] | L. J. Nugent, R. K. Jain, Extravascular diffusion in normal and neoplastic tissues, Cancer Res., 44 (1984), 238–244. |
| [58] | R. K. Jain, Transport of molecules in the tumor interstitium: a review, Cancer Res., 47 (1987), 3039–3051. |
| [59] | L. S. Goodman, Goodman and Gilman's the Pharmacological Basis of Therapeutics, McGraw-Hill New York, 1549 (1996). |
| [60] |
K. M. Laginha, S. Verwoert, G. J. Charrois, T. M. Allen, Determination of doxorubicin levels in whole tumor and tumor nuclei in murine breast cancer tumors, Clin. Cancer Res., 11 (2005), 6944–6949. https://doi.org/10.1158/1078-0432.CCR-05-0343 doi: 10.1158/1078-0432.CCR-05-0343
|
| [61] | A. W. El-Kareh, T. W. Secomb, A mathematical model for comparison of bolus injection, continuous infusion, and liposomal delivery of doxorubicin to tumor cells, Neoplasia, 2 (2000), 325. |
| [62] | A. J. Leu, D. A. Berk, A. Lymboussaki, K. Alitalo, R. K. Jain, Absence of functional lymphatics within a murine sarcoma: a molecular and functional evaluation, Cancer Res., 60 (2000), 4324–4327. |
| [63] |
M. Wu, H. B. Frieboes, S. R. McDougall, M. A. Chaplain, V. Cristini, J. Lowengrub, The effect of interstitial pressure on tumor growth: coupling with the blood and lymphatic vascular systems, J. Theor. Biol., 320 (2013), 131–151. https://doi.org/10.1016/j.jtbi.2012.11.031 doi: 10.1016/j.jtbi.2012.11.031
|
| [64] | G. Powathil, M. Kohandel, M. Milosevic, S. Sivaloganathan, Modeling the spatial distribution of chronic tumor hypoxia: implications for experimental and clinical studies, Comput. Math. Methods Med., 2012 (2012). https: //doi.org/10.1155/2012/410602 |
| [65] |
A. S. Fung, J. Jonkman, I. F. Tannock, Quantitative immunohistochemistry for evaluating the distribution of Ki67 and other biomarkers in tumor sections and use of the method to study repopulation in xenografts after treatment with paclitaxel, Neoplasia, 14 (2012), 324–IN6. https://doi.org/10.1593/neo.12346 doi: 10.1593/neo.12346
|
| [66] |
X. Mao, S. McManaway, J. K. Jaiswal, C. R. Hong, W. R. Wilson, K. O. Hicks, Schedule-dependent potentiation of chemotherapy drugs by the hypoxia-activated prodrug sn30000, Cancer Biol. Ther., 20 (2019), 1258–1269. https://doi.org/10.1080/15384047.2019.1617570 doi: 10.1080/15384047.2019.1617570
|
| [67] |
S. Harris, P. Mistry, C. Freathy, J. Brown, P. Charlton, Antitumour activity of xr5944 in vitro and in vivo in combination with 5-fluorouracil and irinotecan in colon cancer cell lines, Br. J. Cancer, 92 (2005), 722–728. https://doi.org/10.1038/sj.bjc.6602403 doi: 10.1038/sj.bjc.6602403
|
| [68] |
J. M. Saucier, J. Yu, A. Gaikwad, R. L. Coleman, J. K. Wolf, J. A. Smith, Determination of the optimal combination chemotherapy regimen for treatment of platinum-resistant ovarian cancer in nude mouse model, J. Oncol. Pharm. Pract., 13 (2007), 39–45. https://doi.org/10.1177/1078155207077948 doi: 10.1177/1078155207077948
|
| [69] |
M. R. Horsman, L. S. Mortensen, J. B. Petersen, M. Busk, J. Overgaard, Imaging hypoxia to improve radiotherapy outcome, Nat. Rev. Clin. Oncol., 9 (2012), 674–687. https://doi.org/10.1038/nrclinonc.2012.171 doi: 10.1038/nrclinonc.2012.171
|
| [70] | J. Overgaard, Hypoxic modification of radiotherapy in squamous cell carcinoma of the head and neck–a systematic review and meta-analysis, Radiother. Oncol., 100 (2011), 22–32. |
| [71] |
M. Kovacs, D. Hocking, J. Evans, B. Siim, B. Wouters, J. Brown, Cisplatin anti-tumour potentiation by tirapazamine results from a hypoxia-dependent cellular sensitization to cisplatin, Br. J. Cancer, 80 (1999), 1245–1251. https://doi.org/10.1038/sj.bjc.6690492 doi: 10.1038/sj.bjc.6690492
|
| [72] |
X. Zhang, J. W. Wojtkowiak, G. V. Martinez, H. H. Cornnell, C. P. Hart, A. F. Baker, et al., Mr imaging biomarkers to monitor early response to hypoxia-activated prodrug TH-302 in pancreatic cancer xenografts, PloS One, 11 (2016), e0155289. https://doi.org/10.1371/journal.pone.0155289 doi: 10.1371/journal.pone.0155289
|
| [73] |
J. Cárdenas-Rodríguez, Y. Li, J. P. Galons, H. Cornnell, R. J. Gillies, M. D. Pagel, et al., Imaging biomarkers to monitor response to the hypoxia-activated prodrug TH-302 in the miapaca2 flank xenograft model, Magn. Reson. Imaging, 30 (2012), 1002–1009. https://doi.org/10.1016/j.mri.2012.02.015 doi: 10.1016/j.mri.2012.02.015
|
| [74] |
A. Leimgruber, K. Hickson, S. T. Lee, H. K. Gan, L. M. Cher, J. I. Sachinidis, et al., Spatial and quantitative mapping of glycolysis and hypoxia in glioblastoma as a predictor of radiotherapy response and sites of relapse, Eur. J. Nucl. Med. Mol. Imaging, 47 (2020), 1476–1485. https://doi.org/10.1007/s00259-020-04706-0 doi: 10.1007/s00259-020-04706-0
|
| [75] |
B. V. Jardim-Perassi, W. Mu, S. Huang, M. R. Tomaszewski, J. Poleszczuk, M. A. Abdalah, et al., Deep-learning and mr images to target hypoxic habitats with evofosfamide in preclinical models of sarcoma, Theranostics, 11 (2021), 5313. https://doi.org/10.7150/thno.56595 doi: 10.7150/thno.56595
|
| [76] | J. C. Forster, M. J. Douglass, W. M. Harriss-Phillips, E. Bezak, Development of an in silico stochastic 4D model of tumor growth with angiogenesis, Med. Phys., 44 (2017), 1563–1576. |
| [77] | W. M. Harriss-Phillips, E. Bezak, E. Yeoh, The HYP-RT hypoxic tumour radiotherapy algorithm and accelerated repopulation dose per fraction study, Comput. Math. Methods Med., 2012 (2012). https://doi.org/10.1155/2012/363564 |
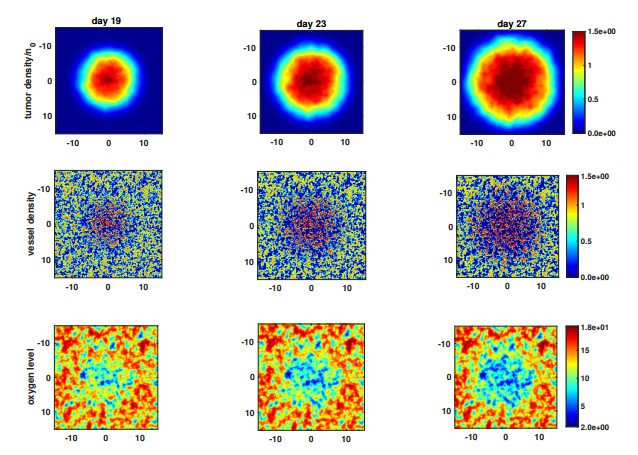
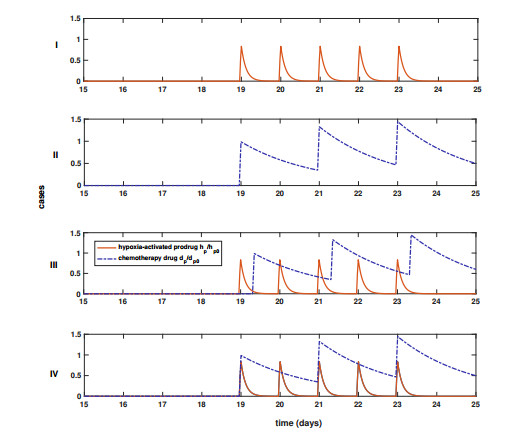
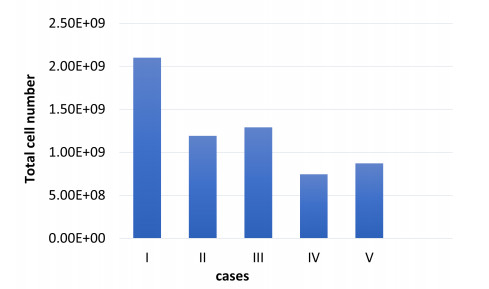
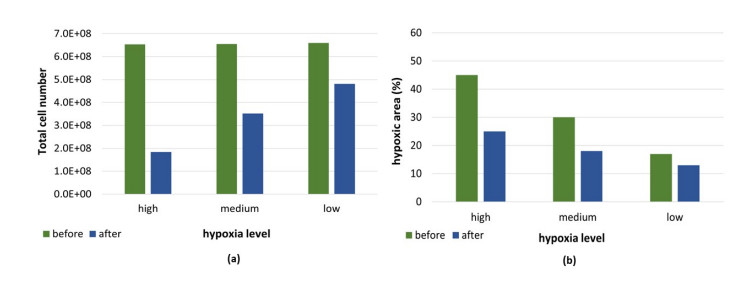
Figures(5) / Tables(3)

Defne Yilmaz, Mert Tuzer, Mehmet Burcin Unlu. Assessing the therapeutic response of tumors to hypoxia-targeted prodrugs with an in silico approach[J]. Mathematical Biosciences and Engineering, 2022, 19(11): 10941-10962. doi: 10.3934/mbe.2022511







 DownLoad:
DownLoad: