Pandemics, such as Covid-19 and AIDS, tend to be highly contagious and have the characteristics of global spread and existence of multiple virus strains. To analyze the competition among different strains, a high dimensional SIR model studying multiple strains' competition in patchy environments is introduced in this work. By introducing the basic reproductive number of different strains, we found global stability conditions of disease-free equilibrium and persistence conditions of the model. The competition exclusion conditions of that model are also given. This work gives some insights into the properties of the multiple strain patchy model and all of the analysis methods used in this work could be used in other related high dimension systems.
Citation: Chentong Li, Jinyan Wang, Jinhu Xu, Yao Rong. The Global dynamics of a SIR model considering competitions among multiple strains in patchy environments[J]. Mathematical Biosciences and Engineering, 2022, 19(5): 4690-4702. doi: 10.3934/mbe.2022218
Pandemics, such as Covid-19 and AIDS, tend to be highly contagious and have the characteristics of global spread and existence of multiple virus strains. To analyze the competition among different strains, a high dimensional SIR model studying multiple strains' competition in patchy environments is introduced in this work. By introducing the basic reproductive number of different strains, we found global stability conditions of disease-free equilibrium and persistence conditions of the model. The competition exclusion conditions of that model are also given. This work gives some insights into the properties of the multiple strain patchy model and all of the analysis methods used in this work could be used in other related high dimension systems.

| [1] | F. Brauer, An introduction to networks in epidemic modeling, Springer, (2008), 133–146. |
| [2] | M. Iannelli, M. Martcheva, F. Milner, Gender-structured population modeling: mathematical methods, numerics, and simulations, Society for Industrial and Applied Mathematics, 2005. https://doi.org/10.1137/1.9780898717488 |
| [3] | J. Cao, X. Qi, H. Zhao, Modeling gene regulation networks using ordinary differential equations, Springer, (2012), 185–197. https://doi.org/10.1007/978-1-61779-400-1_12 |
| [4] |
X. Sun, Y. Xiao, X. Ji, When to lift the lockdown in hubei province during covid-19 epidemic? an insight from a patch model and multiple source data, J. Theor. Biol., 507 (2020), 110469. https://doi.org/10.1016/j.jtbi.2020.110469 doi: 10.1016/j.jtbi.2020.110469

|
| [5] |
D. Bichara, A. Iggidr, G. Sallet, Global analysis of multi-strains sis, sir and msir epidemic models, J. Appl. Math. Comput., 44 (2014), 273–292. https://doi.org/10.1007/s12190-013-0693-x doi: 10.1007/s12190-013-0693-x
|
| [6] |
M. Meehan, D. Cocks, J. Trauer, E. McBryde, Coupled, multi-strain epidemic models of mutating pathogens, Math. biosci., 296 (2018), 82–92. https://doi.org/10.1016/j.mbs.2017.12.006 doi: 10.1016/j.mbs.2017.12.006
|
| [7] |
C. Li, Y. Zhang, Y. Zhou, Spatially antiviral dynamics determines hcv in vivo replication and evolution, J. Theor. Biol., 503 (2020), 110378. https://doi.org/10.1016/j.jtbi.2020.110378 doi: 10.1016/j.jtbi.2020.110378
|
| [8] |
N. Jia, L. Ding, Y. Liu, P. Hu, Global stability and optimal control of epidemic spreading on multiplex networks with nonlinear mutual interaction, Phys. A, 502 (2018), 93–105. https://doi.org/10.1016/j.physa.2018.02.056 doi: 10.1016/j.physa.2018.02.056
|
| [9] |
M. De la Sen, A. Ibeas, S. Alonso-Quesada, R. Nistal, On a sir model in a patchy environment under constant and feedback decentralized controls with asymmetric parameterizations, Symmetry, 11 (2019), 430. https://doi.org/10.3390/sym11030430 doi: 10.3390/sym11030430
|
| [10] |
W. Wang, X. Zhao, An epidemic model in a patchy environment, Math. biosci., 190 (2004), 97–112. https://doi.org/10.1016/j.mbs.2002.11.001 doi: 10.1016/j.mbs.2002.11.001
|
| [11] |
F. Zhang, X. Zhao, A periodic epidemic model in a patchy environment, J. Math. Anal. Appl., 325 (2007), 496–516. https://doi.org/10.1016/j.jmaa.2006.01.085 doi: 10.1016/j.jmaa.2006.01.085
|
| [12] |
M. Li, Z. Shuai, Global stability of an epidemic model in a patchy environment, Can. Appl. Math. Q., 17 (2009), 175–187. https://doi.org/10.1016/j.mbs.2002.11.001 doi: 10.1016/j.mbs.2002.11.001
|
| [13] |
J. Arino, R. Jordan, P. Van den Driessche, Quarantine in a multi-species epidemic model with spatial dynamics, Math. biosci., 206 (2007), 46–60. https://doi.org/10.1016/j.mbs.2005.09.002 doi: 10.1016/j.mbs.2005.09.002
|
| [14] |
M. Marvá, R. de la Parra, J. Poggiale, Approximate aggregation of a two time scales periodic multi-strain sis epidemic model: A patchy environment with fast migrations, Ecol. Complexity, 20 (2012), 34–41. https://doi.org/10.1016/j.ecocom.2011.09.002 doi: 10.1016/j.ecocom.2011.09.002
|
| [15] |
Z. Qiu, Q. Kong, X. Li, M. Martcheva, The vector–host epidemic model with multiple strains in a patchy environment, J. Math. Anal. Appl., 405 (2013), 12–36. https://doi.org/10.1016/j.jmaa.2013.03.042 doi: 10.1016/j.jmaa.2013.03.042
|
| [16] | R. Horn, C. Johnson, Matrix analysis, Cambridge university press, 2012. https://doi.org/10.1017/CBO9780511810817 |
| [17] |
T. Fujimoto, R. Ranade, Two characterizations of inverse-positive matrices: the hawkins-simon condition and the le chatelier-braun principle, Electron. J. Linear Algebra, 11 (2004), 59–65. https://doi.org/10.13001/1081-3810.1122 doi: 10.13001/1081-3810.1122
|
| [18] | W. Walter, Ordinary differential equations, Springer, 1998. |
| [19] | H. Smith, Monotone dynamical systems: an introduction to the theory of competitive and cooperative systems, American Mathematical Society, 2008. https://doi.org/10.1090/surv/041 |
| [20] |
J. Bernal, N. Andrews, C. Gower, E. Gallagher, R. Simmons, S. Thelwall, et al., Effectiveness of covid-19 vaccines against the b. 1.617. 2 (delta) variant, N. Engl. J. Med., 385 (2021), 585–594. https://doi.org/10.1056/NEJMoa2108891 doi: 10.1056/NEJMoa2108891
|
| [21] | Genomic epidemiology of novel coronavirus-global subsampling, 2021. Available from: https://nextstrain.org/ncov/gisaid/global. |
| [22] | C. Li, Y. Zhang, Y. Zhou, Competitive coexistence in a two-strain epidemic model with a periodic infection rate, Discrete Dyn. Nat. Soc., 2020 (2020). https://doi.org/10.1155/2020/7541861 |
| [23] |
X. Zhang, S. Wu, B. Wu, Q. Yang, A. Chen, Y. Li, et al., Sars-cov-2 omicron strain exhibits potent capabilities for immune evasion and viral entrance, Signal Transduct. Target Ther., 6 (2021), 1–3. https://doi.org/10.1038/s41392-021-00852-5 doi: 10.1038/s41392-021-00852-5
|
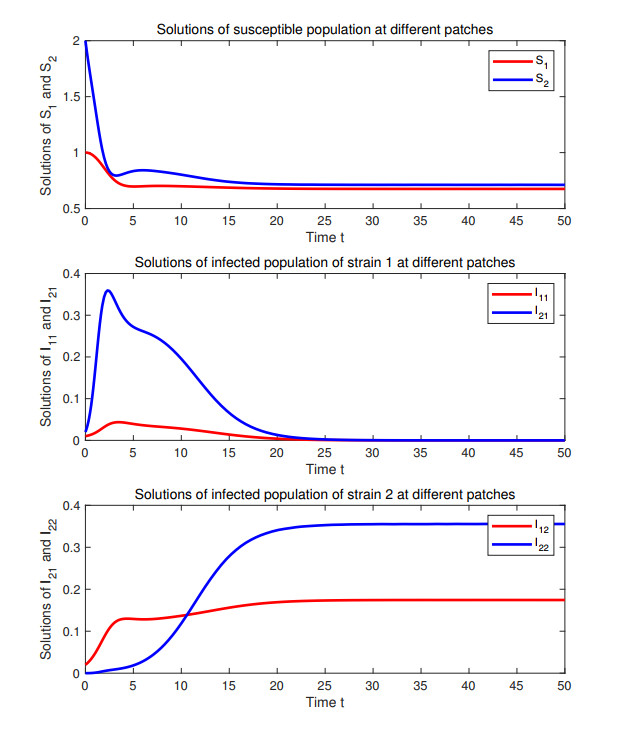


Figures(3)

Chentong Li, Jinyan Wang, Jinhu Xu, Yao Rong. The Global dynamics of a SIR model considering competitions among multiple strains in patchy environments[J]. Mathematical Biosciences and Engineering, 2022, 19(5): 4690-4702. doi: 10.3934/mbe.2022218







 DownLoad:
DownLoad: