We develop a mathematical model, based on a system of ordinary differential equations, to the upshot of farming alertness in crop pest administration, bearing in mind plant biomass, pest, and level of control. Main qualitative analysis of the proposed mathematical model, akin to both pest-free and coexistence equilibrium points and stability analysis, is investigated. We show that all solutions of the model are positive and bounded with initial conditions in a certain significant set. The local stability of pest-free and coexistence equilibria is shown using the Routh–Hurwitz criterion. Moreover, we prove that when a threshold value is less than one, then the pest-free equilibrium is locally asymptotically stable. To get optimum interventions for crop pests, that is, to decrease the number of pests in the crop field, we apply optimal control theory and find the corresponding optimal controls. We establish existence of optimal controls and characterize them using Pontryagin's minimum principle. Finally, we make use of numerical simulations to illustrate the theoretical analysis of the proposed model, with and without control measures.
Citation: Teklebirhan Abraha, Fahad Al Basir, Legesse Lemecha Obsu, Delfim F. M. Torres. Farming awareness based optimum interventions for crop pest control[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 5364-5391. doi: 10.3934/mbe.2021272
We develop a mathematical model, based on a system of ordinary differential equations, to the upshot of farming alertness in crop pest administration, bearing in mind plant biomass, pest, and level of control. Main qualitative analysis of the proposed mathematical model, akin to both pest-free and coexistence equilibrium points and stability analysis, is investigated. We show that all solutions of the model are positive and bounded with initial conditions in a certain significant set. The local stability of pest-free and coexistence equilibria is shown using the Routh–Hurwitz criterion. Moreover, we prove that when a threshold value is less than one, then the pest-free equilibrium is locally asymptotically stable. To get optimum interventions for crop pests, that is, to decrease the number of pests in the crop field, we apply optimal control theory and find the corresponding optimal controls. We establish existence of optimal controls and characterize them using Pontryagin's minimum principle. Finally, we make use of numerical simulations to illustrate the theoretical analysis of the proposed model, with and without control measures.

| [1] |
T. Abraha, F. Al Basir, L. L. Obsu, D. F. M. Torres, Pest control using farming awareness: Impact of time delays and optimal use of biopesticides, Chaos Solitons Fractals, 146 (2021), 110869. doi: 10.1016/j.chaos.2021.110869

|
| [2] |
Y. Li, J. Liang, Models for determining the optimal switching time in chemical control of pest with pesticide resistance, Math. Biosci. Eng., 18 (2021), 471–494. doi: 10.3934/mbe.2021026
|
| [3] |
D. S. Mandal, A. Chekroun, S. Samanta, J. Chattopadhyay, A mathematical study of a crop-pest-natural enemy model with Z-type control, Math. Comput. Simulation, 187 (2021), 468–488. doi: 10.1016/j.matcom.2021.03.014
|
| [4] | M. He, S. Tang, R. A. Cheke, A Holling type II discrete switching host-parasitoid system with a nonlinear threshold policy for integrated pest management, Discrete Dyn. Nat. Soc., 2020 (2020), 1–14. |
| [5] |
C. Li, S. Tang, R. A. Cheke, Complex dynamics and coexistence of period-doubling and period-halving bifurcations in an integrated pest management model with nonlinear impulsive control, Adv. Difference Equations, 2020 (2020), 1–23. doi: 10.1186/s13662-019-2438-0
|
| [6] |
J. Li, Q. Huang, B. Liu, An SI integrated pest management model with pesticide resistance to susceptible pests, Int. J. Biomath., 13 (2020), 2050037. doi: 10.1142/S1793524520500370
|
| [7] |
F. Al Basir, A multi-delay model for pest control with awareness induced interventions–-Hopf bifurcation and optimal control analysis, Int. J. Biomath., 13 (2020), 2050047. doi: 10.1142/S1793524520500473
|
| [8] |
S. E. Naranjo, P. C. Ellsworth, G. B. Frisvold, Economic value of biological control in integrated pest management of managed plant systems, Annu. Rev. Entomol., 60 (2015), 621–645. doi: 10.1146/annurev-ento-010814-021005
|
| [9] | G. A. Khan, S. Muhammad, M. A. Khan, Information regarding agronomic practices and plant protection measures obtained by the farmers through electronic media, J. Anim. Plant Sci., 23 (2013), 647–650. |
| [10] |
J. C. van Lenteren, J. Bale, F. Bigler, H. M. T. Hokkanen, A. J. M. Loomans, Assessing risks of releasing exotic biological control agents of arthropod pests, Annu. Rev. Entomol., 51 (2006), 609–634. doi: 10.1146/annurev.ento.51.110104.151129
|
| [11] | X. Yang, F. Wang, L. Meng, W. Zhang, L. Fan V. Geissen, C. J. Ritsema, Farmer and retailer knowledge and awareness of the risks from pesticide use: A case study in the Wei River catchment, China, Sci. Total Environ., 497 (2014), 172–179. |
| [12] |
F. Al Basir, A. Banerjee, S. Ray, Role of farming awareness in crop pest management–-a mathematical model, J. Theoret. Biol., 461 (2019), 59–67. doi: 10.1016/j.jtbi.2018.10.043
|
| [13] |
F. Le Bellec, A. Rajaud, H. Ozier-Lafontaine, C. Bockstaller, E. Malezieux, Evidence for farmers' active involvement in co-designing citrus cropping systems using an improved participatory method, Agron. Sustain. Dev., 32 (2012), 703–714. doi: 10.1007/s13593-011-0070-9
|
| [14] |
F. Al Basir, E. Venturino, P. K. Roy, Effects of awareness program for controlling mosaic disease in Jatropha curcas plantations, Math. Methods Appl. Sci., 40 (2017), 2441–2453. doi: 10.1002/mma.4149
|
| [15] |
J. Chowdhury, F. Al Basir, Y. Takeuchi, M. Ghosh, P. K. Roy, A mathematical model for pest management in Jatropha curcas with integrated pesticides–-an optimal control approach, Ecol. Complexity, 37 (2019), 24–31. doi: 10.1016/j.ecocom.2018.12.004
|
| [16] | M. Junusov, Optimal control problems connected with agrocenotic models, Dokl. Akad. Nauk Tadžik. SSR, 21 (1978), 10–14. |
| [17] |
A. Molter, M. Rafikov, Optimal control in agroecosystems using the method of state-dependent Riccati equations, TEMA Tend. Mat. Apl. Comput., 12 (2011), 221–232. doi: 10.5540/tema.2011.012.03.0221
|
| [18] | C. J. Silva, D. F. M. Torres, E. Venturino, Optimal spraying in biological control of pests, Math. Model. Nat. Phenom., 12 (2017), 51–64. |
| [19] |
F. Al Basir, S. Ray, Impact of farming awareness based roguing, insecticide spraying and optimal control on the dynamics of mosaic disease, Ric. Mat., 69 (2020), 393–412. doi: 10.1007/s11587-020-00522-8
|
| [20] |
S. Pathak, A. Maiti, Pest control using virus as control agent: a mathematical model, Nonlinear Anal. Model. Control, 17 (2012), 67–90. doi: 10.15388/NA.17.1.14079
|
| [21] | W. H. Fleming, R. W. Rishel, Deterministic and stochastic optimal control, Springer-Verlag, Berlin, 1975. |
| [22] | S. Lenhart, J. T. Workman, Optimal control applied to biological models, Chapman & Hall/CRC, Boca Raton, FL, 2007. |
| [23] | M. I. Kamien, N. L. Schwartz, Dynamic optimization, second edition, Advanced Textbooks in Economics, 31, North-Holland Publishing Co., Amsterdam, 1991. |
| [24] |
S. Jana, T. K. Kar, A mathematical study of a prey-predator model in relevance to pest control, Nonlinear Dynam., 74 (2013), 667–683. doi: 10.1007/s11071-013-0996-3
|
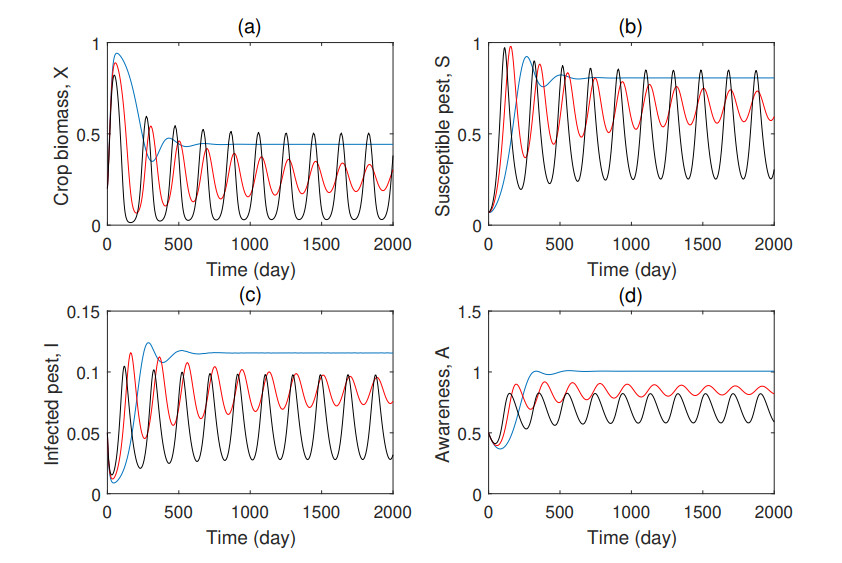
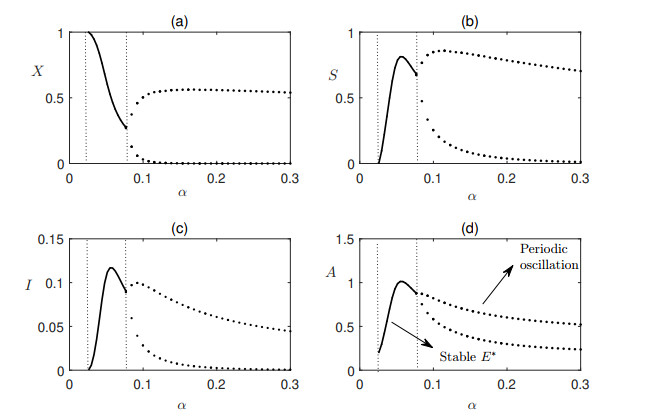
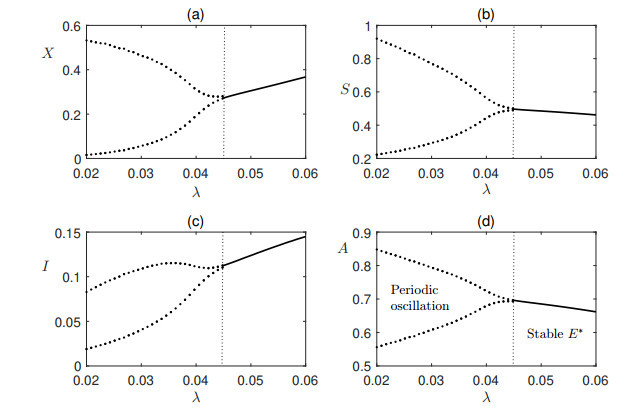
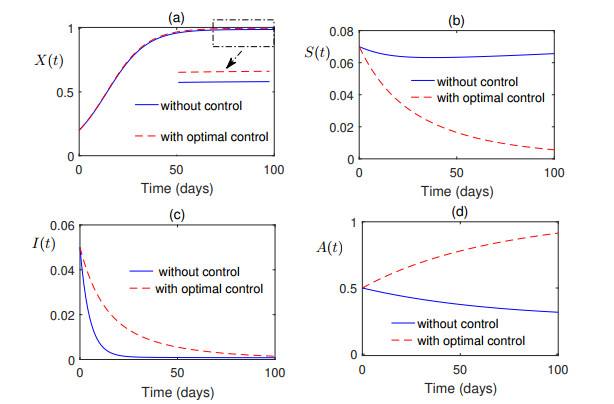
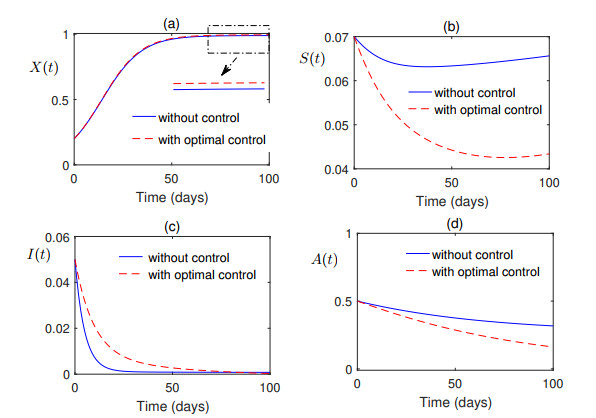
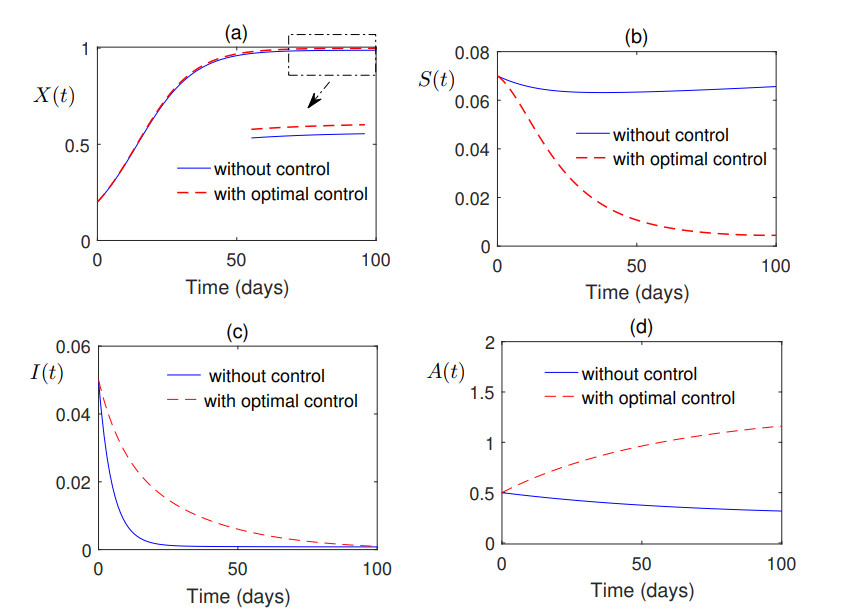
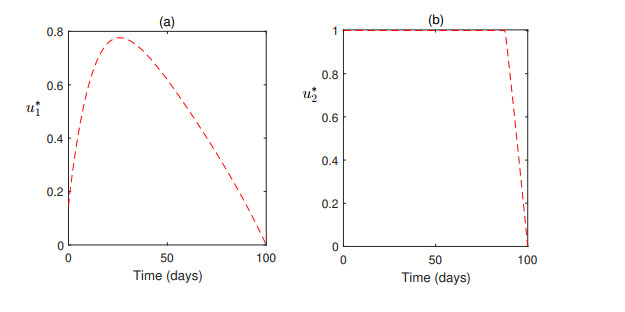
Figures(7) / Tables(1)

Teklebirhan Abraha, Fahad Al Basir, Legesse Lemecha Obsu, Delfim F. M. Torres. Farming awareness based optimum interventions for crop pest control[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 5364-5391. doi: 10.3934/mbe.2021272







 DownLoad:
DownLoad: