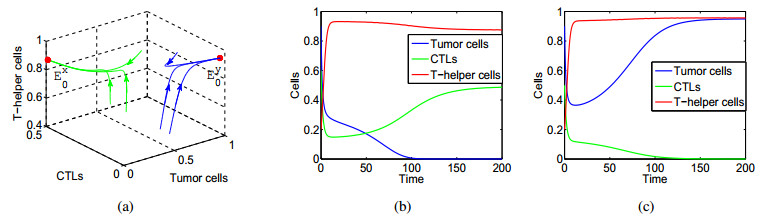
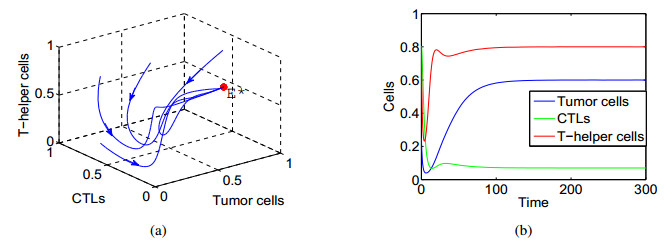
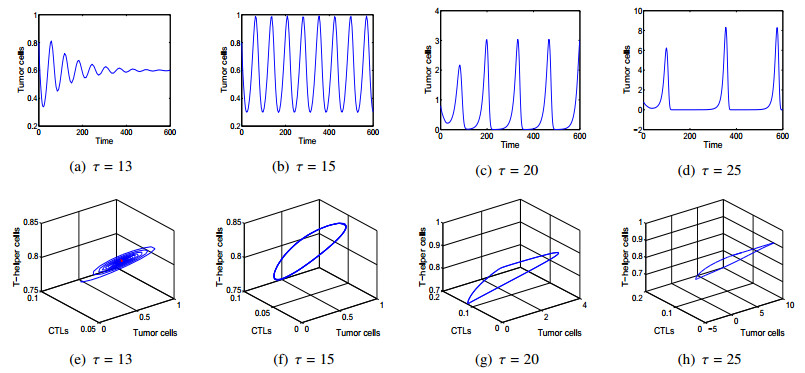
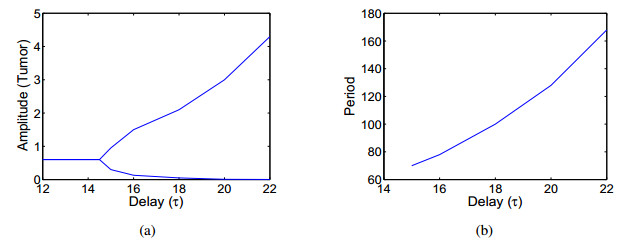
In this paper, a three-dimensional nonlinear delay differential system including Tumour cells, cytotoxic-T lymphocytes, T-helper cells is constructed to investigate the effects of intrinsic recruitment delay and chemotherapy. It is found that the introduction of chemotherapy and time delay can generate richer dynamics in tumor-immune system. In particular, there exists bistable phenomenon and the tumour cells would be cleared if the effect of chemotherapy on depletion of the tumour cells is strong enough or the side effect of chemotherapy on the hunting predator cells is under a threshold. It is also shown that a branch of stable periodic solutions bifurcates from the coexistence equilibrium when the intrinsic recruitment delay of tumor crosses the threshold which is new mechanism, which can help understand the short-term oscillations in tumour sizes as well as long-term tumour relapse. Numerical simulations are presented to illustrate that larger intrinsic recruitment delay of tumor leads to larger amplitude and longer period of the bifurcated periodic solution, which indicates that there exists longer relapse time and then contributes to the control of tumour growth.
Citation: Qingfeng Tang, Guohong Zhang. Stability and Hopf bifurcations in a competitive tumour-immune system with intrinsic recruitment delay and chemotherapy[J]. Mathematical Biosciences and Engineering, 2021, 18(3): 1941-1965. doi: 10.3934/mbe.2021101
In this paper, a three-dimensional nonlinear delay differential system including Tumour cells, cytotoxic-T lymphocytes, T-helper cells is constructed to investigate the effects of intrinsic recruitment delay and chemotherapy. It is found that the introduction of chemotherapy and time delay can generate richer dynamics in tumor-immune system. In particular, there exists bistable phenomenon and the tumour cells would be cleared if the effect of chemotherapy on depletion of the tumour cells is strong enough or the side effect of chemotherapy on the hunting predator cells is under a threshold. It is also shown that a branch of stable periodic solutions bifurcates from the coexistence equilibrium when the intrinsic recruitment delay of tumor crosses the threshold which is new mechanism, which can help understand the short-term oscillations in tumour sizes as well as long-term tumour relapse. Numerical simulations are presented to illustrate that larger intrinsic recruitment delay of tumor leads to larger amplitude and longer period of the bifurcated periodic solution, which indicates that there exists longer relapse time and then contributes to the control of tumour growth.

| [1] |
L. M. F. Merlo, J. W. Pepper, B. J. Reid, C. C. Maley, Cancer as an evolutionary and ecological process, Nat. Rev. Cancer, 6 (2006), 924–935. doi: 10.1038/nrc2013

|
| [2] | A. Jemal, F. Bray, M. M. Center, Global cancer statistics, Ca. Cancer. J. Clin., 61 (2011), 69–90. |
| [3] |
R. R. Sarkar, S. Banerjee, Cancer self remission and tumor stability-a stochastic approach, Math. Biosci., 196 (2005), 65–81. doi: 10.1016/j.mbs.2005.04.001
|
| [4] | K. Subhas, J. J. Nieto, Mathematical modeling of tumor-immune competitive system, considering the role of time delay, Appl. Math. Comput., 340 (2019), 180–205. |
| [5] |
D. Kirschner, J. Panetta, Modelling immunotherapy of the tumor-immune interaction, J. Math. Biol., 37 (1998), 235–252. doi: 10.1007/s002850050127
|
| [6] | L. G. de Pillis, A. Radunskaya, The dynamics of an optimally controlled tumor model: A case study, Math. Comput. Model., 37 (2003), 1221–1244. |
| [7] |
N. Tsur, Y. Kogana, M. Rehmc, Z. Agur, Response of patients with melanoma to immune checkpoint blockade-insights gleaned from analysis of a new mathematical mechanistic model, J. Theor. Biol., 485 (2020), 110033. doi: 10.1016/j.jtbi.2019.110033
|
| [8] |
M. A. Owen, J. A. Sherratt, Mathematical modelling macrophage dynamics in tumors, Math. Model. Meth. Appl. Sci., 9 (1999), 513–539. doi: 10.1142/S0218202599000270
|
| [9] | J. L. Yu, S. R. J. Jang, A mathematical model of tumor-immune interactions with an immune checkpoint inhibitor, Appl. Math. Comput., 362 (2019), 124532. |
| [10] |
R. A. Ku-Carrillo, S. E. Delgadillo, B. M. Chen-Charpentier, A mathematical model for the effect of obesity on cancer growth and on the immune system response, Appl. Math. Model., 40 (2016), 4908–4920. doi: 10.1016/j.apm.2015.12.018
|
| [11] |
N. Bellomo, L. Preziosi, Modelling and mathematical problems related to tumor evolution and its interaction with the immune system, Math. Comput. Model., 32 (2000), 413–452. doi: 10.1016/S0895-7177(00)00143-6
|
| [12] |
V. A. Kuznetsov, I. A. Makalkin, M. A. Taylor, A. S. Perelson, Nonlinear dynamics of immunogenic tumors: Parameter estimation and global bifurcation analysis, Bull. Math. Biol., 56 (1994), 295–321. doi: 10.1016/S0092-8240(05)80260-5
|
| [13] |
C. Letellier, F. Denis, L. A. Aguirre, What can be learned from a chaotic cancer model?, J. Theor. Biol., 322 (2013), 7–16. doi: 10.1016/j.jtbi.2013.01.003
|
| [14] |
A. Zazoua, W. D. Wang, Analysis of mathematical model of prostate cancer with androgen deprivation therapy, Commun. Nonlinear. Sci. Numer. Simul., 66 (2019), 41–60. doi: 10.1016/j.cnsns.2018.06.004
|
| [15] |
L. Y. Pang, Z. Zhao, X. Y. Song, Cost-effectiveness analysis of optimal strategy for tumor treatment, Chaos Solitons Fractals, 87 (2016), 293–301. doi: 10.1016/j.chaos.2016.03.032
|
| [16] |
S. Khajanchi, S. Banerjee, Quantifying the role of immunotherapeutic drug T11 target structure in progression of malignant gliomas: Mathematical modeling and dynamical perspective, Math. Biosci., 289 (2017), 69–77. doi: 10.1016/j.mbs.2017.04.006
|
| [17] |
S. Banerjee, S. Khajanchi, S. Chaudhur, A mathematical model to elucidate brain tumor abrogation by immunotherapy with T11 target structure, PLoS One, 10 (2015), 1–21. doi: 10.1371/journal.pone.0116884
|
| [18] | J. Arciero, T. Jackson, D. Kirschner, A mathematical model of tumor-immune evasion and siRNA treatment, Discret. Contin. Dyn. Syst. Ser. B., 4 (2004), 39–58. |
| [19] |
L. G. depillis, A. Eladdadi, A. E. Radunskaya, Modeling cancer-immune responses to therapy, J. Pharmacokinet. Pharmacodyn., 41 (2014), 461–0478. doi: 10.1007/s10928-014-9386-9
|
| [20] |
M. Itik, M. U. Salamci, S. P. Banks, Optimal control of drug therapy in cancer treatment, Nonlinear Anal., 71 (2009), 1473–1486. doi: 10.1016/j.na.2009.01.214
|
| [21] |
X. D. Liu, Q. Z. Li, J. X. Pan, A deterministic and stochastic model for the system dynamics of tumor-immune responses to chemotherapy, Phys. A, 500 (2018), 162–176. doi: 10.1016/j.physa.2018.02.118
|
| [22] |
L. G. depillis, W. Gu, K. R. Fister, T. Head, K. Maples, A. Murugan, et al., Chemotherapy for tumors: An analysis of the dynamics and a study of quadratic and linear optimal controls, Math. Biosci., 209 (2007), 292–315. doi: 10.1016/j.mbs.2006.05.003
|
| [23] |
P. Rokhforoz, A. A. Jamshidi, N. N. Sarvestani, Adaptive robust control of cancer chemotherapy with extended Kalman filter observer, Inform. Med. Unlock., 8 (2017), 1–7. doi: 10.1016/j.imu.2017.03.002
|
| [24] |
L. G. de Pillis, W. Gu, A. E. Radunskaya, Mixed immunotherapy and chemotherapy of tumors: modeling, applications and biological interpretations, J. Theor. Biol., 238 (2006), 841–862. doi: 10.1016/j.jtbi.2005.06.037
|
| [25] |
R. Eftimie, J. J. Gillard, D. A. Cantrell, Mathematical models for immunology: Current state of the art and future research directions, Bull. Math. Biol., 78 (2016), 2091–2134. doi: 10.1007/s11538-016-0214-9
|
| [26] |
M. Villasana, A. Radunskaya, A delay differential equation model for tumor growth, J. Math. Biol., 47 (2003), 270–294. doi: 10.1007/s00285-003-0211-0
|
| [27] |
Y. Radouane, Hopf bifurcation in differential equations with delay for tumor-immune system competition model, SIAM J. Appl. Math., 67 (2007), 1693–1703. doi: 10.1137/060657947
|
| [28] | F. A. Rihan, D. H. Abdel Rahman, S. Lakshmanan, A. S. Alkhajeh, A time delay model of tumour-immune system interactions: Global dynamics, parameter estimation, sensitivity analysis, Appl. Math. Comput., 232 (2014), 606–623. |
| [29] |
M. Yu, Y. P. Dong, Y. Takeuchi, Dual role of delay effects in a tumour-immune system, J. Biol. Dyn., 11 (2017), 334–347. doi: 10.1080/17513758.2016.1231347
|
| [30] | S. Khajanchi, S. Banerjee, Stability and bifurcation analysis of delay induced tumor immune interaction model, Appl. Math. Comput., 248 (2014), 652–671. |
| [31] |
S. Khajanchi, Bifurcation analysis of a delayed mathematical model for tumor growth, Chaos Solitons Fractals, 77 (2015), 264–276. doi: 10.1016/j.chaos.2015.06.001
|
| [32] |
S. Khajanchi, S. Banerjee, Influence of multiple delays in brain tumor and immune system interaction with T11 target structure as a potent stimulator, Math. Biosci., 302 (2018), 116–130. doi: 10.1016/j.mbs.2018.06.001
|
| [33] |
M. J. Piotrowska, M. Bodnar, Influence of distributed delays on the dynamics of a generalized immune system cancerous cells interactions model, Commun. Nonlinear. Sci. Numer. Simul., 54 (2018), 389–415. doi: 10.1016/j.cnsns.2017.06.003
|
| [34] |
S. Banerjee, R. R. Sarkar, Delay-induced model for tumor-immune interaction and control of malignant tumor growth, BioSystems, 91 (2008), 268–288. doi: 10.1016/j.biosystems.2007.10.002
|
| [35] |
A. E. Gohary, Chaos and optimal control of cancer self-remission and tumor system steady states, Chaos Solitons Fractals, 37 (2008), 1305–1316. doi: 10.1016/j.chaos.2006.10.060
|
| [36] |
H. Mayer, K. Zaenker, U. an der Heiden, A basic mathematical model of the immune response, Chaos, 5 (1995), 155–161. doi: 10.1063/1.166098
|
| [37] |
H. M. Byrne, The effect of time delay on the dynamics of avascular tumor growth, Math. Biosci., 144 (1997), 83–117. doi: 10.1016/S0025-5564(97)00023-0
|
| [38] | P. Bi, S. G. Ruan, Bifurcations in delay differential equations and applications to tumor and immune system interaction models, SIAM J. Appl. Dyn., 12 (2014), 1847–1888. |
| [39] | S. G. Ruan, Nonlinear dynamics in tumor-immune system interaction models with delays, Discret. Contin. Dyn. Syst. Ser. B., 26 (2021), 541–602. |
| [40] |
F. A. Rihan, Sensitivity analysis of dynamic systems with time lags, J. Comput. Appl. Math., 151 (2003), 445–462. doi: 10.1016/S0377-0427(02)00659-3
|
| [41] |
K. Gopalsamy, Nonoscillation in a delay-logistic equation, Quart. Appl. Math., 43 (1985), 189–197. doi: 10.1090/qam/793526
|
| [42] |
D. Mukherjee, P. C. Bhakta, A. B. Roy, Uniform persistence in Kolmogorov models with convex growth functions, Nonlinear Anal., 34 (1998), 427–432. doi: 10.1016/S0362-546X(97)00605-6
|
| [43] | J. J. Wei, S. G. Ruan, Stability and bifurcation in a neural network model with two delays, Phys. D, 130 (1998), 255–272. |
| [44] | B. D. Hassard, N. D. Kazarinoff, Y. H. Wan, Theory and Application of Hopf Bifurcation, London Mathematical Society Lecture Note Series, Cambridge University Press, 1981. |
Figures(5) / Tables(2)

Qingfeng Tang, Guohong Zhang. Stability and Hopf bifurcations in a competitive tumour-immune system with intrinsic recruitment delay and chemotherapy[J]. Mathematical Biosciences and Engineering, 2021, 18(3): 1941-1965. doi: 10.3934/mbe.2021101







 DownLoad:
DownLoad: