Citation: Maoxiang Wang, Fenglan Hu, Meng Xu, Zhipeng Qiu. Keep, break and breakout in food chains with two and three species[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 817-836. doi: 10.3934/mbe.2021043

| [1] | N. J. Gotelli, A. M. Ellison, Food-web models predict species abundances in response to habitat change, PLoS Biol., 4 (2006), 1869-1873. |
| [2] |
S. B. Hsu, T. W. Hwang, Y. Kuang, A Ratio-dependent food chain model and its applications to control, Math. Biosci., 181 (2003), 55-83. doi: 10.1016/S0025-5564(02)00127-X

|
| [3] |
C. H. Chiu, S. B. Hsu, Extinction of top-predator in a three-level food-chain model, J. Math. Biol., 37 (1998), 372-380. doi: 10.1007/s002850050134
|
| [4] |
K. R. Crooks, M. E. Soulé, Mesopredator release and avifaunal extinctions in a fragmented system, Nature, 400 (1999), 563-566. doi: 10.1038/23028
|
| [5] |
J. R. Britton, Introduced parasites in food webs: new species, shifting structures?, Trends Ecol. Evol., 28 (2013), 93-99. doi: 10.1016/j.tree.2012.08.020
|
| [6] | A. J. Lotka, Elements of Physical Biology, Williams and Willkins, Baltimore, 1925. |
| [7] |
V. Volterra, Variations and fluctuations of the number of individuals in animal species living together, ICES. J. Mar. Sci., 3 (1928), 3-51. doi: 10.1093/icesjms/3.1.3
|
| [8] |
R. Arditi, L.R. Ginzburg, Coupling in predator-prey dynamics: ratio-dependence, J. Theor. Biol., 139 (1989), 311-326. doi: 10.1016/S0022-5193(89)80211-5
|
| [9] |
A. Fenton, M. Spencer, D. J. S. Montagnes, Parameterising variable assimilation efficiency in predator-prey models, Oikos, 119 (2010), 1000-1010. doi: 10.1111/j.1600-0706.2009.17875.x
|
| [10] |
J. Li, D. J. Montagnes, Restructuring fundamental predator-prey models by recognizing prey-dependent conversion efficiency and mortality rates, Protist, 166 (2015), 211-223. doi: 10.1016/j.protis.2015.02.003
|
| [11] | T. K. Kar, A. Ghorai, A. Batabyal, Global dynamics and bifurcation of a tri-trophic food chain model, World J. Model. Simul., 8 (2012), 66-80. |
| [12] |
E. E. Joshua, E. T. Akpan, C. E. Madubueze, Hopf-bifurcation limit cycles of an extended Rosenzweig-MacArthur model, J. Math. Res., 8 (2016), 22. doi: 10.5539/jmr.v8n3p22
|
| [13] |
B. Deng, Food chain chaos due to junction-fold point, Chaos, 11 (2001), 514. doi: 10.1063/1.1396340
|
| [14] |
B. Deng, G. Hines, Food chain chaos due to transcritical point, Chaos, 13 (2003), 578. doi: 10.1063/1.1576531
|
| [15] |
D. M. Post, The long and short of food-chain length, Trends Ecol. Evol., 17 (2002), 269. doi: 10.1016/S0169-5347(02)02455-2
|
| [16] | M. Kondon, K. Ninomiya, Food-chain length and adaptive foraging, P. Roy. Soc. B: Biol. Sci., 276 (2009), 3113-3121. |
| [17] |
M. L. Pace, J. J. Cole, S. R. Carpenter, J. F. Kitchell, Trophic cascades revealed in diverse ecosystems, Trends Ecol. Evol., 14 (1999), 483-488. doi: 10.1016/S0169-5347(99)01723-1
|
| [18] |
L. Persson, Trophic cascades: Abiding heterogeneity and the trophic level concept at the end of the road, Oikos, 85 (1999), 385-397. doi: 10.2307/3546688
|
| [19] |
L. Oksanen, T. Oksanen, The logic and realism of the hypothesis of exploitation ecosystems, Am. Nat., 155 (2000), 703-723. doi: 10.1086/303354
|
| [20] | J. Hofbauer, K. Sigmund, Evolutionary games and population dynamics, Cambridge University Press, Cambridge, 1998. |
| [21] | C. P. Rocaa, J. A. Cuesta, A. Sanchez, Evolutionary game theory: Temporal and spatial effects beyond replicator dynamics, Phys. Life Rev., 6 (2000), 208. |
| [22] |
D. Melese, S. Gakkhar, Pattern formation in tri-trophic ratio-dependent food chain model, Appl. Math., 2 (2011), 1507-1514. doi: 10.4236/am.2011.212213
|
| [23] |
M. C. Cross, P. C. Hohenberg, Pattern formation out of equilibrium, Rev. Mod. Phys., 65 (1993) 851. doi: 10.1103/RevModPhys.65.851
|
| [24] |
W. van Saarloos, P. C. Hohenberg, Fronts, pulses, sources, and sinks in generalized complex Ginzburg-Landau equations, Phys. D, 56 (1992), 303. doi: 10.1016/0167-2789(92)90175-M
|
| [25] | S. Amadae, Prisoner's Dilemma, Prisoners of Reason, Cambridge University Press, New York, 2016. |
| [26] |
M. H. Vainstein, J. J. Arenzon, Spatial social dilemmas: Dilution, mobility, and grouping effects with imitation dynamics, Phys. A, 394 (2014), 145. doi: 10.1016/j.physa.2013.09.032
|
| [27] |
M. X. Wang, Y. Q. Ma, P. Y. Lai, Regulatory effects on the population dynamics and wave propagation in a cell lineage model, J. Theor. Biol., 393 (2016), 105-117. doi: 10.1016/j.jtbi.2015.12.035
|
| [28] |
M. X. Wang, P. Y. Lai, Population dynamics and wave propagation in Lotka-Volterra system with spatial diffusion, Phys. Rev. E, 86 (2012), 051908. doi: 10.1103/PhysRevE.86.051908
|
| [29] |
M. X. Wang, Y. Ma, Population evolution in mutualistic Lotka-Volterra system with spatial diffusion, Phys. A, 395 (2014), 228-235. doi: 10.1016/j.physa.2013.10.019
|
| [30] |
H. Q. Zhu, M. X. Wang, P. Y. Lai, General two-species interacting Lotka-Volterra system: Population dynamics and wave propagation, Phys. Rev. E, 97 (2018), 052413. doi: 10.1103/PhysRevE.97.052413
|
| [31] |
H. Q. Zhu, M. X. Wang, F. L. Hu, Interaction and coexistence with self-regulating species, Phys. A, 502 (2018), 447. doi: 10.1016/j.physa.2018.02.082
|
| [32] |
M. L. Rosenzweig, R. H. MacArthur, Graphical representation and stability conditions of predator-prey interactions, Am. Nat., 97 (1963), 209-223; doi: 10.1086/282272
|
| [33] |
J. P. Chen, H. D. Zhang, The qualitative analysis of two species predator-prey model with Holling's type Ⅲ functional response, Appl. Math. Mech., 7 (1986), 77-86. doi: 10.1007/BF01896254
|
| [34] |
C. S. Holling, The components of predation as revealed by a study of small mammal predation of the European pine sawfly, Can. Entomol., 91 (1959), 293-320. doi: 10.4039/Ent91293-5
|
| [35] |
C. S. Holling, Some characteristics of simple types of predation and parasitism, Can. Entomol., 91 (1959), 385-398. doi: 10.4039/Ent91385-7
|
| [36] | C. S. Holling, The functional response of predators to prey density and its role in mimicry and population regulation, Mem. Entomol. Soc. Can., 45 (1959), 5-60. |
| [37] |
R. Peng, J. Shi, M. Wang, Stationary pattern of a ratio-dependent food chain model with diffusion, SIAM J. Appl. Math., 67 (2007), 1479-1503. doi: 10.1137/05064624X
|
| [38] |
R. R. Vance, The Effect of Dispersal on Population Stability in One-Species, Discrete-Space Population Growth Models, Am. Nat., 123 (1984), 230. doi: 10.1086/284199
|
| [39] |
L. J. S. Allen, Persistence and extinction in Lotka-Volterra reaction-diffusion equations, Math. Biosci., 65 (1983), 1. doi: 10.1016/0025-5564(83)90068-8
|
| [40] |
Y. Takeuchi, Diffusion effect on stability of Lotka-Volterra models, Bull. Math. Biol., 48 (1986), 585-601. doi: 10.1016/S0092-8240(86)90009-1
|
| [41] |
A. Hastings, Global stability in Lotka-Volterra systems with diffusion, J. Math. Biol., 6 (1978) 163-168. doi: 10.1007/BF02450786
|
| [42] | R. A. Fisher, The advance of advantageous genes, Ann. Eugenics, 7 (1937), 353. |
| [43] | A. Kolmogorov, I. Petrovskii, N. Piscounov, Selected Works of A. N. Kolmogorov, Vol. 2, I Math. Appl., 1991 (1991), 25. |
| [44] |
M. M. Tang and P. C. Fife, Propagating fronts for competing species equations with diffusion, Arch. Ration. Mech. Anal., 73 (1980), 69. doi: 10.1007/BF00283257
|
| [45] |
J. I. Kanel, L. Zhou, Existence of wave front solutions and estimates of wave speed for a competition-diffusion system, Nonlinear Anal. TMA, 27 (1996), 579. doi: 10.1016/0362-546X(95)00221-G
|
| [46] |
E. Barbera, G. Consolo, G. Valenti, A two or three compartments hyperbolic reaction-diffusion model for the aquatic food chain, Math. Biosci. Eng., 12 (2015), 451. doi: 10.3934/mbe.2015.12.451
|
| [47] |
C. Erica, J. E. Paullet, J. P. Previte, Z. Walls, A Lotka- Volterra three-species food chain, Math. Mag., 75 (2002), 243-255. doi: 10.1080/0025570X.2002.11953139
|
| [48] |
O. De Feo, S. Rinaldi, Yield and dynamics of tri-trophic food chains, Am. Nat., 150 (1997), 328-345. doi: 10.1086/286068
|
| [49] |
C. Borrvall, B. Ebenman, T. Jonsson, Biodiversity lessens the risk of cascading extinction in model food webs, Ecol. Lett., 3 (2000), 131-136. doi: 10.1046/j.1461-0248.2000.00130.x
|
| [50] |
C. H. Chiu, S. B. Hsu, Extinction of top-predator in a three-level food-chain model, J. Math. Biol., 37 (1998), 372-380. doi: 10.1007/s002850050134
|
| [51] |
K. R. Crooks, M. E. Soulé, Mesopredator release and avifaunal extinctions in a fragmented system, Nature, 400 (1999), 563-566. doi: 10.1038/23028
|
| [52] |
K. S. McCann, J. B. Rasmussen, J. Umbanhowar, The dynamics of spatially coupled food webs, Eco. Lett., 8 (2005), 513-523. doi: 10.1111/j.1461-0248.2005.00742.x
|
| [53] |
M. X. Wang, Y. J. Li, P. Y. Lai, C. K. Chan, Model on cell movement, growth, differentiation and de-differentiation: Reaction-diffusion equation and wave propagation, Eur. Phys. J. E, 36 (2013), 65. doi: 10.1140/epje/i2013-13065-4
|
| [54] |
C, Liu, X. Fu, L. Liu, X. Ren, C. K. L. Chau, S. Li, et al., Sequential establishment of stripe patterns in an expanding cell population, Science, 334 (2011), 238. doi: 10.1126/science.1209042
|
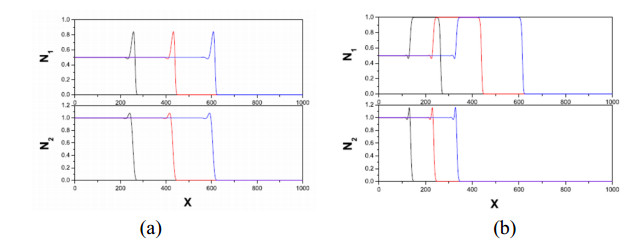
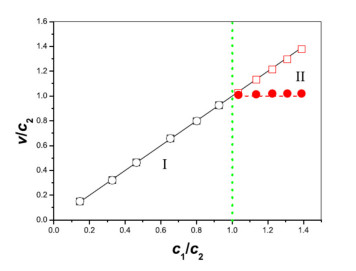
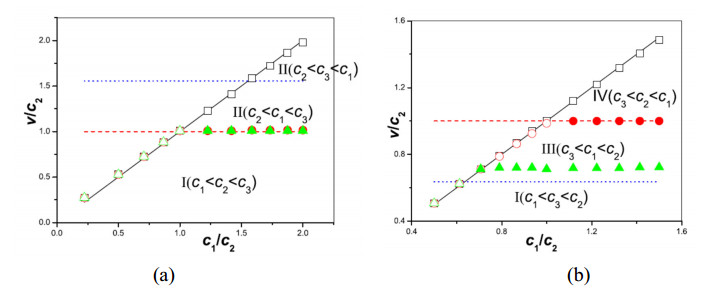
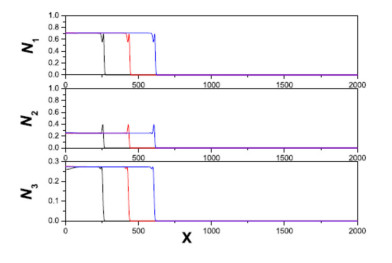
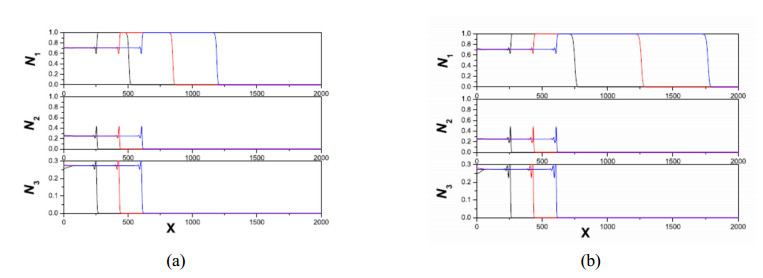
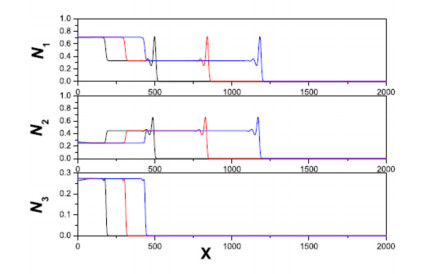
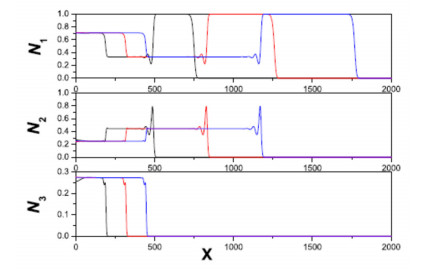
Figures(11)

Maoxiang Wang, Fenglan Hu, Meng Xu, Zhipeng Qiu. Keep, break and breakout in food chains with two and three species[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 817-836. doi: 10.3934/mbe.2021043







 DownLoad:
DownLoad: