Citation: Jian-Di Li, Gang Chen, Mei Wu, Yu Huang, Wei Tang. Downregulation of CDC14B in 5218 breast cancer patients: A novel prognosticator for triple-negative breast cancer[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 8152-8181. doi: 10.3934/mbe.2020414

| [1] |
F. Bray, J. Ferlay, I. Soerjomataram, R. L. Siegel, L. A. Torre, A. Jemal, Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries, CA Cancer J. Clin., 68 (2018), 394-424. doi: 10.3322/caac.21492

|
| [2] |
R. L. Siegel, K. D. Miller, A. Jemal, Cancer statistics, CA Cancer J. Clin., 70 (2020), 7-30. doi: 10.3322/caac.21590
|
| [3] |
C. Larsson, A. Ehinger, S. Winslow, K. Leandersson, M. Klintman, L. Dahl, et al., Prognostic implications of the expression levels of different immunoglobulin heavy chain-encoding RNAs in early breast cancer, NPJ Breast Cancer, 6 (2020), 28. doi: 10.1038/s41523-020-0170-2
|
| [4] |
M. Alsaleem, M. S. Toss, C. Joseph, M. Aleskandarany, S. Kurozumi, I. Alshankyty, et al., The molecular mechanisms underlying reduced E-cadherin expression in invasive ductal carcinoma of the breast: high throughput analysis of large cohorts, Mod. Pathol., 32 (2019), 967-976. doi: 10.1038/s41379-019-0209-9
|
| [5] |
S. Yoon, H. S. Won, K. Kang, K. Qiu, W. J. Park, Y. H. Ko, Hormone Receptor-Status Prediction in Breast Cancer Using Gene Expression Profiles and Their Macroscopic Landscape, Cancers, 12 (2020), 1165. doi: 10.3390/cancers12051165
|
| [6] |
A. Nasir, M. M. H. Bullo, Z. Ahmed, A. Imtiaz, E. Yaqoob, M. Jadoon, et al., Nutrigenomics: Epigenetics and cancer prevention: A comprehensive review, Crit. Rev. Food Sci. Nutr., 60 (2020), 1375-1387. doi: 10.1080/10408398.2019.1571480
|
| [7] |
D. R. Chen, W. C. Hsieh, Y. L. Liao, K. J. Lin, Y. F. Wang, P. H. Lin, Imbalances in the disposition of estrogen and naphthalene in breast cancer patients: a potential biomarker of breast cancer risk, Sci. Rep., 10 (2020), 11773. doi: 10.1038/s41598-020-68814-5
|
| [8] |
D. Ma, Y. Z. Jiang, Y. Xiao, M. D. Xie, S. Zhao, X. Jin, et al., Integrated molecular profiling of young and elderly patients with triple-negative breast cancer indicates different biological bases and clinical management strategies, Cancer, 126 (2020), 3209-3218. doi: 10.1002/cncr.32922
|
| [9] |
J. Chen, M. K. Haanpaa, J. J. Gruber, N. Jager, J. M. Ford, M. P. Snyder, High-Resolution Bisulfite-Sequencing of Peripheral Blood DNA Methylation in Early-Onset and Familial Risk Breast Cancer Patients, Clin. Cancer Res., 25 (2019), 5301-5314. doi: 10.1158/1078-0432.CCR-18-2423
|
| [10] |
I. V. Ulasov, A. V. Borovjagin, P. Timashev, M. Cristofanili, D. R. Welch, KISS1 in breast cancer progression and autophagy, Cancer Metastasis Rev., 38 (2019), 493-506. doi: 10.1007/s10555-019-09814-4
|
| [11] |
M. S. M. Issac, E. Yousef, M. R. Tahir, L. A. Gaboury, MCM2, MCM4, and MCM6 in Breast Cancer: Clinical Utility in Diagnosis and Prognosis, Neoplasia (N. Y., NY, U. S.), 21 (2019), 1015-1035. doi: 10.1016/j.neo.2019.07.011
|
| [12] |
Q. Wu, B. Li, Z. Li, J. Li, S. Sun, S. Sun, Cancer-associated adipocytes: key players in breast cancer progression, J. Hematol. Oncol., 12 (2019), 95. doi: 10.1186/s13045-019-0778-6
|
| [13] |
R. Eyre, D. G. Alferez, A. Santiago-Gomez, K. Spence, J. C. McConnell, C. Hart, et al., Microenvironmental IL1β promotes breast cancer metastatic colonisation in the bone via activation of Wnt signalling, Nat. Commun., 10 (2019), 5016. doi: 10.1038/s41467-019-12807-0
|
| [14] |
K. Pandey, H. J. An, S. K. Kim, S. A. Lee, S. Kim, S. M. Lim, et al., Molecular mechanisms of resistance to CDK4/6 inhibitors in breast cancer: A review, Int. J. Cancer, 145 (2019), 1179-1188. doi: 10.1002/ijc.32020
|
| [15] |
K. Goutsouliak, J. Veeraraghavan, V. Sethunath, C. De Angelis, C. K. Osborne, M. F. Rimawi, et al., Towards personalized treatment for early stage HER2-positive breast cancer, Nat. Rev. Clin. Oncol., 17 (2020), 233-250. doi: 10.1038/s41571-019-0299-9
|
| [16] | Collaborative Group on Hormonal Factors in Breast Cancer, Type and timing of menopausal hormone therapy and breast cancer risk: individual participant meta-analysis of the worldwide epidemiological evidence, Lancet, 394 (2019), 1159-1168. |
| [17] |
G. E. Naoum, L. Salama, A. Ho, N. K. Horick, O. Oladeru, M. Abouegylah, et al., The Impact of Chest Wall Boost on Reconstruction Complications and Local Control in Patients Treated for Breast Cancer, Int. J. Radiat. Oncol., Biol., Phys., 105 (2019), 155-164. doi: 10.1016/j.ijrobp.2019.04.027
|
| [18] | A. Darlix, G. Louvel, J. Fraisse, W. Jacot, E. Brain, M. Debled, et al., Impact of breast cancer molecular subtypes on the incidence, kinetics and prognosis of central nervous system metastases in a large multicentre real-life cohort, Br. J. Cancer, 121 (2019), 1-10. |
| [19] |
S. M. Tolaney, A. M. Wardley, S. Zambelli, J. F. Hilton, T. A. Troso-Sandoval, F. Ricci, et al., Abemaciclib plus trastuzumab with or without fulvestrant versus trastuzumab plus standard-of-care chemotherapy in women with hormone receptor-positive, HER2-positive advanced breast cancer (monarcHER): a randomised, open-label, phase 2 trial, Lancet Oncol., 21 (2020), 763-775. doi: 10.1016/S1470-2045(20)30112-1
|
| [20] | J. Li, K. Yu, D. Pang, C. Wang, J. Jiang, S. Yang, et al., Adjuvant Capecitabine With Docetaxel and Cyclophosphamide Plus Epirubicin for Triple-Negative Breast Cancer (CBCSG010): An Open-Label, Randomized, Multicenter, Phase III Trial, J. Clin. Oncol., 38 (2020), 1774-1784. |
| [21] |
P. Sun, D. Zhang, H. Huang, Y. Yu, Z. Yang, Y. Niu, et al., MicroRNA-1225-5p acts as a tumor-suppressor in laryngeal cancer via targeting CDC14B, Biol. Chem., 400 (2019), 237-246. doi: 10.1515/hsz-2018-0265
|
| [22] |
S. Ovejero, P. Ayala, M. Malumbres, F. X. Pimentel-Muinos, A. Bueno, M. P. Sacristan, Biochemical analyses reveal amino acid residues critical for cell cycle-dependent phosphorylation of human Cdc14A phosphatase by cyclin-dependent kinase 1, Sci. Rep., 8 (2018), 11871. doi: 10.1038/s41598-018-30253-8
|
| [23] |
M. Dietachmayr, A. Rathakrishnan, O. Karpiuk, F. von Zweydorf, T. Engleitner, V. Fernandez-Saiz, et al., Antagonistic activities of CDC14B and CDK1 on USP9X regulate WT1-dependent mitotic transcription and survival, Nat. Commun., 11 (2020), 1268. doi: 10.1038/s41467-020-15059-5
|
| [24] |
H. Lin, K. Ha, G. Lu, X. Fang, R. Cheng, Q. Zuo, et al., Cdc14A and Cdc14B Redundantly Regulate DNA Double-Strand Break Repair, Mol. Cell. Biol., 35 (2015), 3657-3668. doi: 10.1128/MCB.00233-15
|
| [25] | Y. Wang, J. Tian, C. Huang, J. Ma, G. Hu, Y. Chen, et al., P53 suppresses SENP3 phosphorylation to mediate G2 checkpoint, Cell Discovery, 6 (2020), 21. |
| [26] |
Y. Kim, J. W. Choi, J. H. Lee, Y. S. Kim, Loss of CDC14B expression in clear cell renal cell carcinoma: meta-analysis of microarray data sets, Am. J. Clin. Pathol., 141 (2014), 551-558. doi: 10.1309/AJCP4PE4JPSRGBQS
|
| [27] |
D. M. Kokkinakis, X. Liu, S. Chada, M. M. Ahmed, M. M. Shareef, U. K. Singha, et al., Modulation of gene expression in human central nervous system tumors under methionine deprivation-induced stress, Cancer Res., 64 (2004), 7513-7525. doi: 10.1158/0008-5472.CAN-04-0592
|
| [28] |
D. F. Calvisi, S. Ladu, F. Pinna, M. Frau, M. L. Tomasi, M. Sini, et al., SKP2 and CKS1 promote degradation of cell cycle regulators and are associated with hepatocellular carcinoma prognosis, Gastroenterology, 137 (2009), 1816-1826. doi: 10.1053/j.gastro.2009.08.005
|
| [29] |
B. Wu, X. Chen, J. Wang, X. Qing, Z. Wang, X. Ding, et al., Separation and characterization of extracellular vesicles from human plasma by asymmetrical flow field-flow fractionation, Anal. Chim. Acta, 1127 (2020), 234-245. doi: 10.1016/j.aca.2020.06.071
|
| [30] |
L. Tu, R. Guan, H. Yang, Y. Zhou, W. Hong, L. Ma, et al., Assessment of the expression of the immune checkpoint molecules PD-1, CTLA4, TIM-3 and LAG-3 across different cancers in relation to treatment response, tumor-infiltrating immune cells and survival, Int. J. Cancer, 147 (2020), 423-439. doi: 10.1002/ijc.32785
|
| [31] |
M. J. Goldman, J. Zhang, N. A. Fonseca, I. Cortes-Ciriano, Q. Xiang, B. Craft, et al., A user guide for the online exploration and visualization of PCAWG data, Nat. Commun., 11 (2020), 3400. doi: 10.1038/s41467-020-16785-6
|
| [32] |
E. Vagia, D. Mahalingam, M. Cristofanilli, The Landscape of Targeted Therapies in TNBC, Cancers, 12 (2020), 916. doi: 10.3390/cancers12040916
|
| [33] |
R. Yang, L. Xing, M. Wang, H. Chi, L. Zhang, J. Chen, Comprehensive Analysis of Differentially Expressed Profiles of lncRNAs/mRNAs and miRNAs with Associated ceRNA Networks in Triple-Negative Breast Cancer, Cell. Physiol. Biochem., 50 (2018), 473-488. doi: 10.1159/000494162
|
| [34] |
J. Wang, Y. Su, Y. Tian, Y. Ding, X. Wang, Characterization of DNA hydroxymethylation profile in cervical cancer, Artif. Cells, Nanomed., Biotechnol., 47 (2019), 2706-2714. doi: 10.1080/21691401.2019.1634578
|
| [35] |
M. N. H. Luijten, J. X. T. Lee, K. C. Crasta, Mutational game changer: Chromothripsis and its emerging relevance to cancer, Mutat. Res., 777 (2018), 29-51. doi: 10.1016/j.mrrev.2018.06.004
|
| [36] |
D. A. Quigley, H. X. Dang, S. G. Zhao, P. Lloyd, R. Aggarwal, J. J. Alumkal, et al., Genomic Hallmarks and Structural Variation in Metastatic Prostate Cancer, Cell, 174 (2018), 758-769. doi: 10.1016/j.cell.2018.06.039
|
| [37] | A. Marcozzi, F. Pellestor, W. P. Kloosterman, The Genomic Characteristics and Origin of Chromothripsis, Chromothripsis, Humana Press, New York, NY, 1769 (2018), 3-9. |
| [38] | The ICGC/TCGA Pan-Cancer Analysis of Whole Genomes Consortium, Pan-cancer analysis of whole genomes, Nature, 578 (2020), 82-93. |
| [39] |
N. Voronina, J. K. L. Wong, D. Hubschmann, M. Hlevnjak, S. Uhrig, C. E. Heilig, et al., The landscape of chromothripsis across adult cancer types, Nat. Commun., 11 (2020), 2320. doi: 10.1038/s41467-020-16134-7
|
| [40] | Z. Li, X. Zhang, C. Hou, Y. Zhou, J. Chen, H. Cai, et al., Comprehensive identification and characterization of somatic copy number alterations in triple‑negative breast cancer, Int. J. Oncol., 56 (2020), 522-530. |
| [41] |
F. Hong, S. Pan, Y. Guo, P. Xu, Y. Zhai, PPARs as nuclear receptors for nutrient and energy metabolism, Molecules, 24 (2019), 2545. doi: 10.3390/molecules24142545
|
| [42] |
W. H. Chang, A. G. Lai, An integrative pan-cancer investigation reveals common genetic and transcriptional alterations of AMPK pathway genes as important predictors of clinical outcomes across major cancer types, BMC cancer, 20 (2020), 773. doi: 10.1186/s12885-020-07286-2
|
| [43] |
Y. Liu, J. K. Colby, X. Zuo, J. Jaoude, D. Wei, I. Shureiqi, The role of PPAR-δ in metabolism, inflammation, and cancer: Many characters of a critical transcription factor, Int. J. Mol. Sci., 19 (2018), 3339. doi: 10.3390/ijms19113339
|
| [44] |
W. H. Chang, A. G. Lai, The pan-cancer mutational landscape of the PPAR pathway reveals universal patterns of dysregulated metabolism and interactions with tumor immunity and hypoxia, Ann. N. Y. Acad. Sci., 1448 (2019), 65-82. doi: 10.1111/nyas.14170
|
| [45] |
G. Sultan, S. Zubair, I. A. Tayubi, H. U. Dahms, I. H. Madar, Towards the early detection of ductal carcinoma (a common type of breast cancer) using biomarkers linked to the PPAR(γ) signaling pathway, Bioinformation, 15 (2019), 799-805. doi: 10.6026/97320630015799
|
| [46] |
Y. P. Hwang, S. S. Won, S. W. Jin, G. H. Lee, T. H. Pham, J. H. Choi, et al., WY-14643 regulates CYP1B1 expression through peroxisome proliferator-activated receptor α-mediated signaling in human breast cancer cells, Int. J. Mol. Sci., 20 (2019), 5928. doi: 10.3390/ijms20235928
|
| [47] |
L. Ponnusamy, S. R. Natarajan, K. Thangaraj, R. Manoharan, Therapeutic aspects of AMPK in breast cancer: Progress, challenges, and future directions, Biochim. Biophys. Acta, Rev. Cancer, 1874 (2020), 188379. doi: 10.1016/j.bbcan.2020.188379
|
| [48] |
S. C. Kwong, A. H. A. Jamil, A. Rhodes, N. A. Taib, I. Chung, Metabolic role of fatty acid binding protein 7 in mediating triple-negative breast cancer cell death via PPAR-α signaling, J. Lipid Res., 60 (2019), 1807-1817. doi: 10.1194/jlr.M092379
|
| [49] |
M. K. Apaya, P. W. Hsiao, Y. C. Yang, L. F. Shyur, Deregulating the CYP2C19/epoxy-eicosatrienoic acid-associated FABP4/FABP5 signaling network as a therapeutic approach for metastatic triple-negative breast cancer, Cancers, 12 (2020), 199. doi: 10.3390/cancers12010199
|
| [50] |
H. Liu, H. Sun, B. Zhang, S. Liu, S. Deng, Z. Weng, et al., F-FDG PET imaging for monitoring the early anti-tumor effect of albendazole on triple-negative breast cancer, Breast cancer, 27 (2020), 372-380. doi: 10.1007/s12282-019-01027-5
|
| [51] |
H. Fang, G. Du, Q. Wu, R. Liu, C. Chen, J. Feng, HDAC inhibitors induce proline dehydrogenase (POX) transcription and anti-apoptotic autophagy in triple negative breast cancer, Acta Biochim. Biophys. Sin., 51 (2019), 1064-1070. doi: 10.1093/abbs/gmz097
|
| [52] | D. Zhou, X. Zhao, M. Yu, Y. Xu, C. Fu, K. Zheng, et al., Anti-migration and anti-invasion effects of 2-hydroxy-6-tridecylbenzoic acid is associated with the enhancement of CYP1B1 expression through activating the AMPK signaling pathway in triple-negative breast cancer cells, Nat. Prod. Res., 2020 (2020), 1-5. |
| [53] |
N. Bhuiyan, P. Singh, S. M. Harden, S. K. Mama, Rural physical activity interventions in the United States: a systematic review and RE-AIM evaluation, Int. J. Behav. Nutr. Phys. Act., 16 (2019), 140. doi: 10.1186/s12966-019-0903-5
|
| [54] |
G. Pasello, A. Pavan, I. Attili, A. Bortolami, L. Bonanno, J. Menis, et al., Real world data in the era of Immune Checkpoint Inhibitors (ICIs): Increasing evidence and future applications in lung cancer, Cancer Treat Rev., 87 (2020), 102031. doi: 10.1016/j.ctrv.2020.102031
|
| [55] |
I. Meisingset, O. Vasseljen, N. K. Vøllestad, H. S. Robinson, A. Woodhouse, K. B. Engebretsen, et al., Novel approach towards musculoskeletal phenotypes, Eur. J. Pain, 24 (2020), 921-932. doi: 10.1002/ejp.1541
|
| [56] |
C. Lee, K. J. Fernandez, S. Alexandrou, C. M. Sergio, N. Deng, S. Rogers, et al., Cyclin E2 Promotes Whole Genome Doubling in Breast Cancer, Cancers, 12 (2020), 2268. doi: 10.3390/cancers12082268
|
 MBE-17-06-414-Supplementary.pdf MBE-17-06-414-Supplementary.pdf |
 |

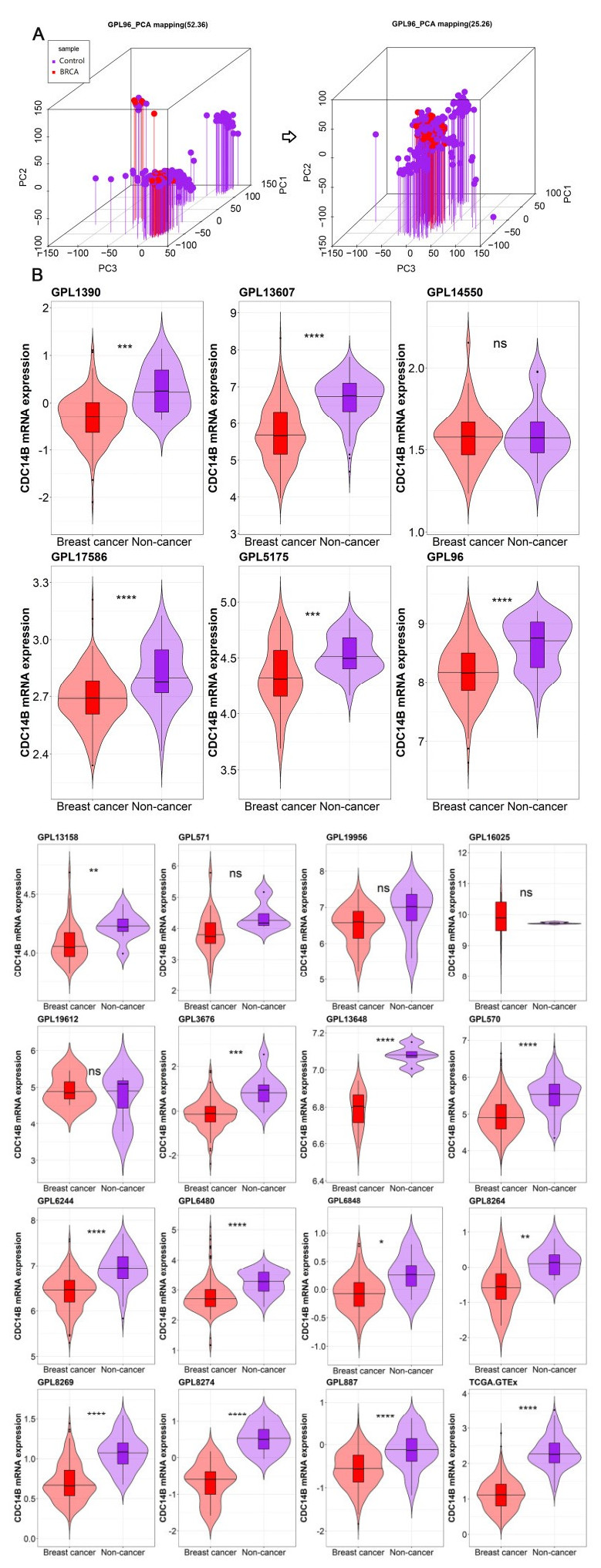
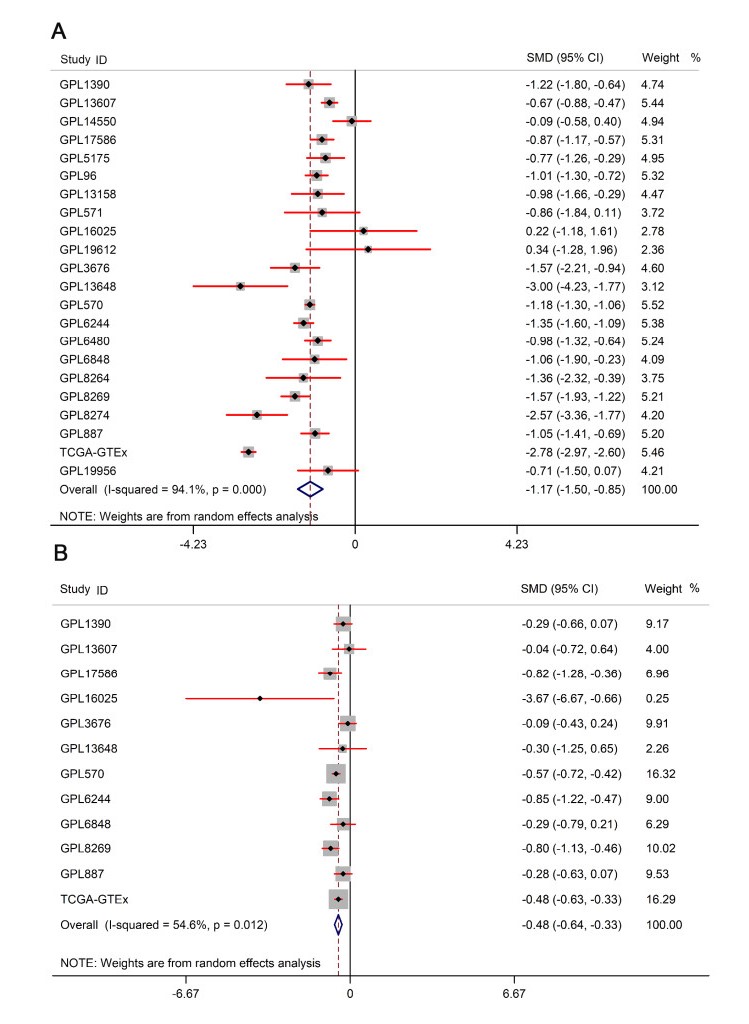
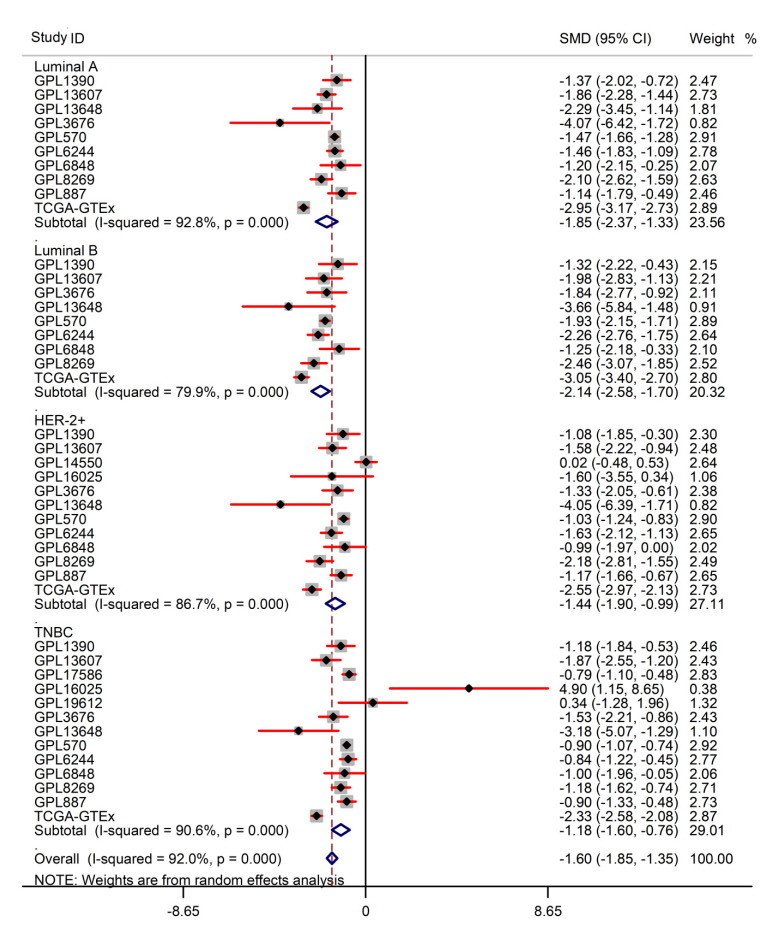
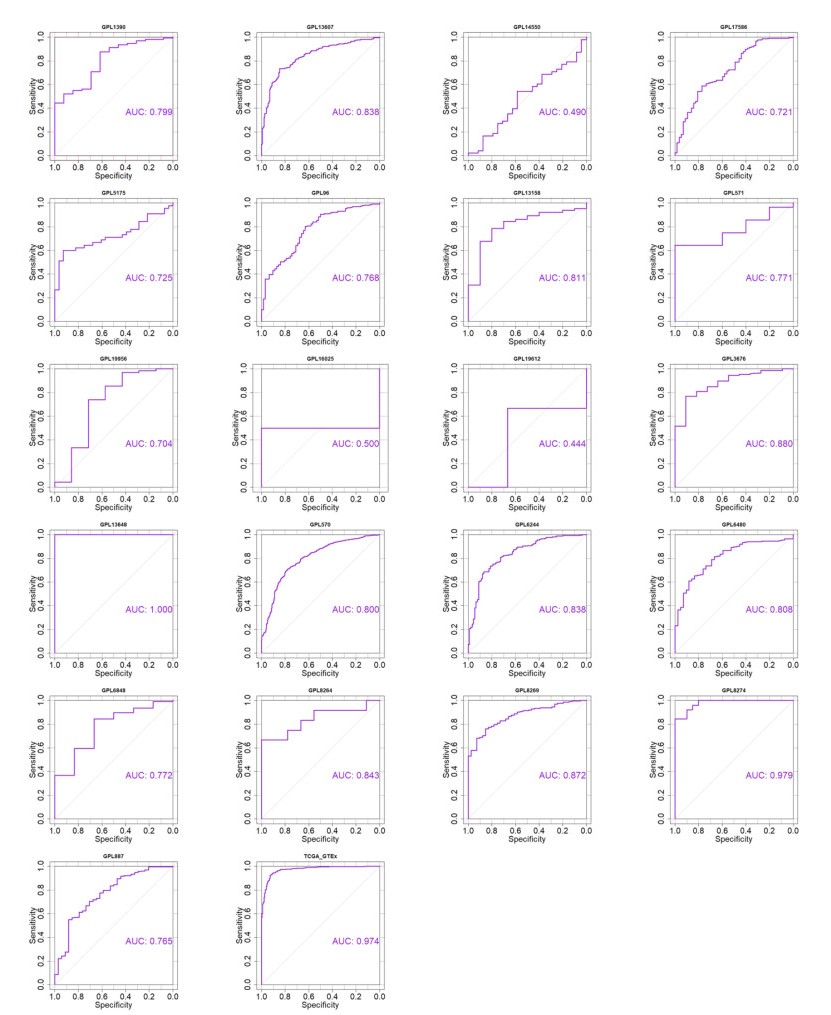

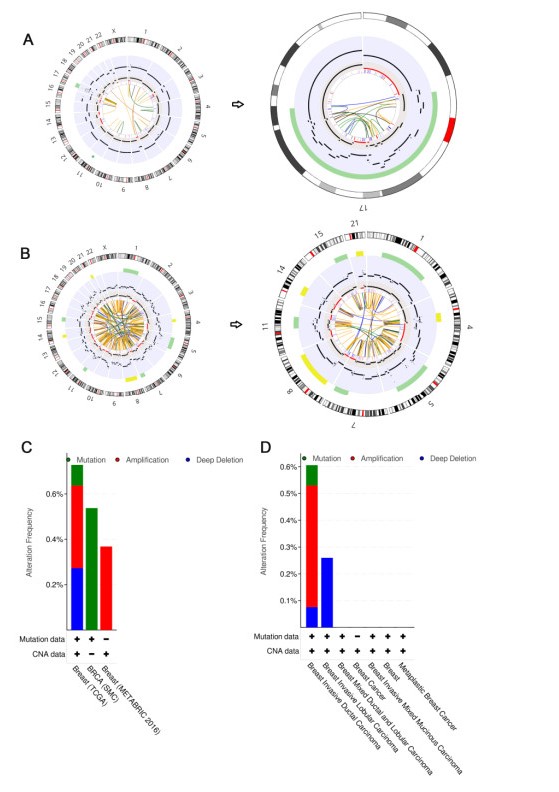
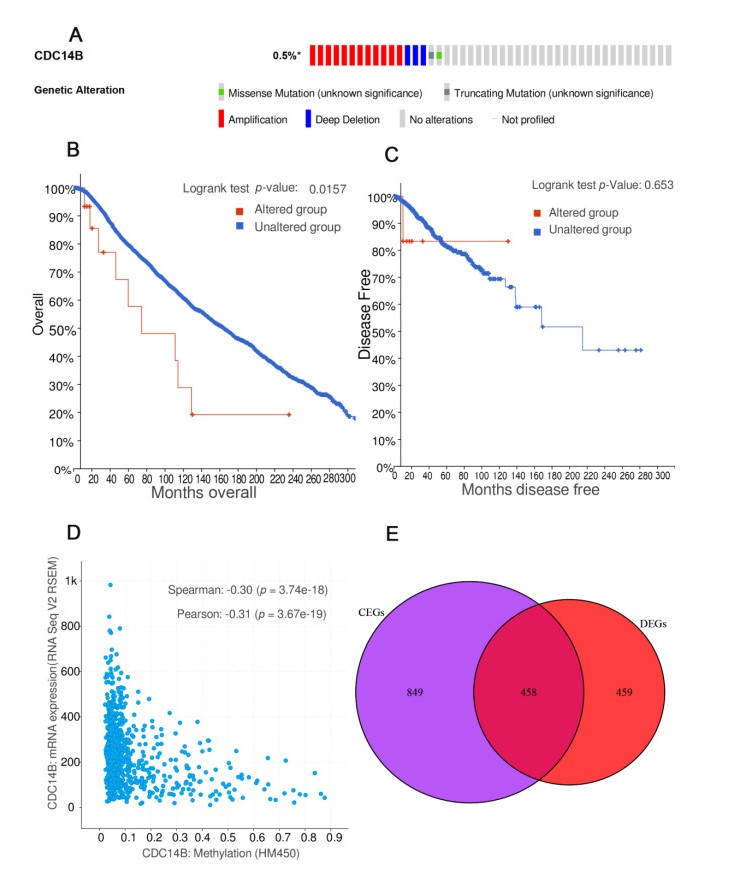
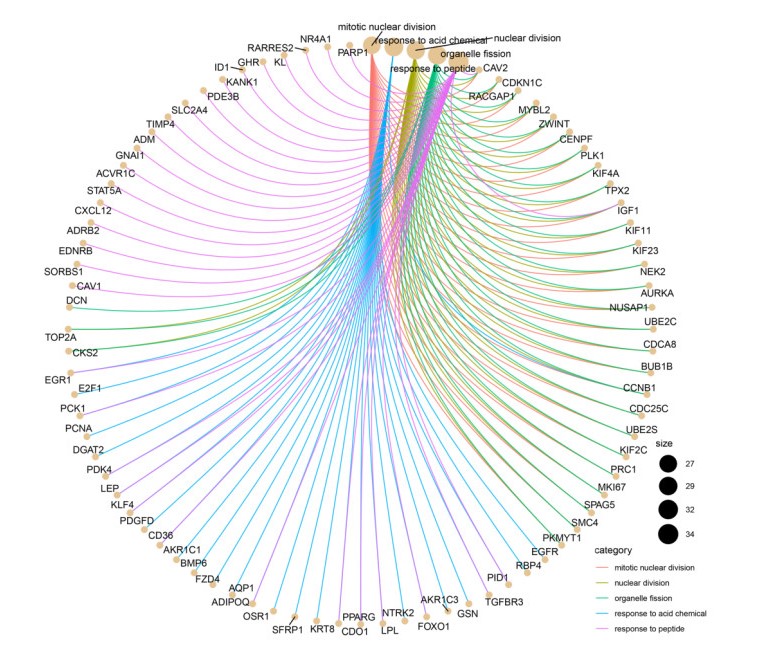

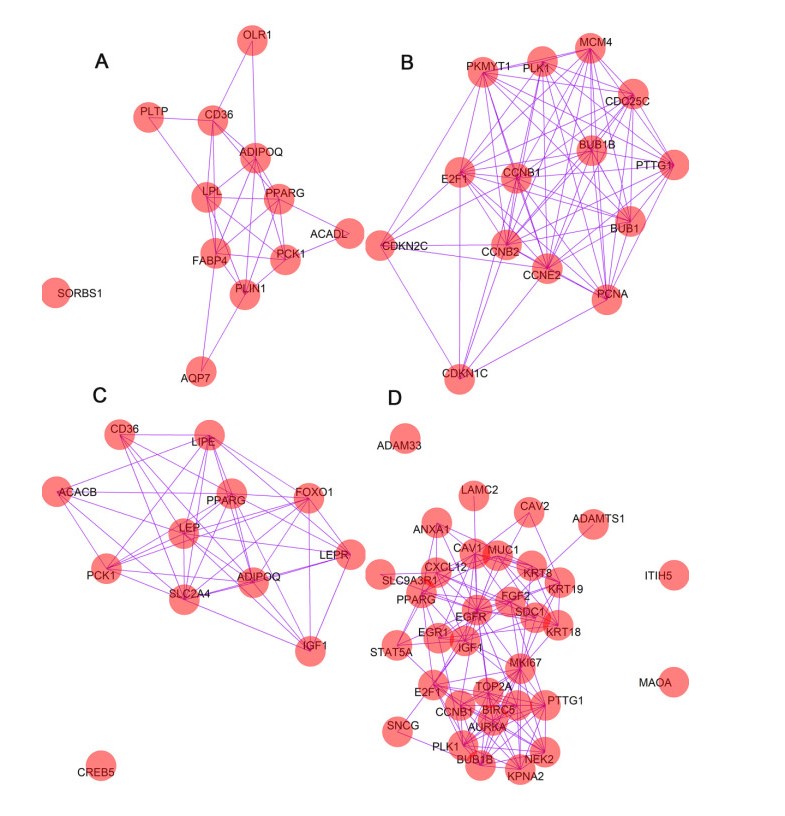

Figures(13) / Tables(1)

Jian-Di Li, Gang Chen, Mei Wu, Yu Huang, Wei Tang. Downregulation of CDC14B in 5218 breast cancer patients: A novel prognosticator for triple-negative breast cancer[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 8152-8181. doi: 10.3934/mbe.2020414







 DownLoad:
DownLoad: