Citation: Matthew D. Johnston, Bruce Pell. A dynamical framework for modeling fear of infection and frustration with social distancing in COVID-19 spread[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7892-7915. doi: 10.3934/mbe.2020401

| [1] | Worldometers.info, Covid-19 coronavirus pandemic, available from: https://www.worldometers.info/coronavirus/. Accessed: 29 Sept, 2020. |
| [2] |
A. J. Kucharski, T. W. Russell, C. Diamond, Y. Liu, J. Edmunds, S. Funk, et al., Early dynamics of transmission and control of COVID-19: A mathematical modelling study, Lancet Infect. Dis., 20 (2020), P553-558. doi: 10.1016/S1473-3099(20)30144-4

|
| [3] |
S. Zhang, M. Y. Diao, W. Yuc, L. Pei, Z. Lin, D. Chen, et al., Estimation of the reproductive number of novel coronavirus (COVID-19) and the probable outbreak size on the diamond princess cruise ship: A data-driven analysis, Int. J. Infect. Dis., 93 (2020), 201-204. doi: 10.1016/j.ijid.2020.02.033
|
| [4] |
Z. Zhuang, S. Zhao, Q. Lin, P. Cao, Y. Lou, L. Yang, et al., Preliminary estimates of the reproduction number of the coronavirus disease (COVID-19) outbreak in Republic of Korea and Italy by 5 March 2020, Int. J. Infect. Dis., 95 (2020), 308-310. doi: 10.1016/j.ijid.2020.04.044
|
| [5] | M. Lv, X. Luo, J. Estill, Y. Liu, M. Ren, J. Wang, et al., Coronavirus disease (COVID-19): a scoping review, Euro. Surveil., 25 (2020), 2000125. |
| [6] | M. V. Barbarossa, J. Fuhrmann, J. Heidecke, H. V. Varma, N. Castelletti, J. H. Meinke, et al., A first study on the impact of current and future control measures on the spread of COVID-19 in Germany, medRxiv, (2020). Available from: https://www.medrxiv.org/content/10.1101/2020.04.08.20056630v1. |
| [7] |
A. Bouchnita, A. Jebrane, A hybrid multi-scale model of COVID-19 transmission dynamics to assess the potential of non-pharmaceutical interventions, Chaos Soliton Fract., 138 (2020), 109941. doi: 10.1016/j.chaos.2020.109941
|
| [8] | S. L. Chang, N. Harding, C. Zachreson, O. M. Cliff, M. Prokopenko, Modelling transmission and control of the COVID-19 pandemic in Australia, arXiv, (2020). Available from: arXiv: 2003.10218v2. |
| [9] | R. Djidjou-Demassea, Y. Michalakisa, M. Choisya, M. T. Sofonea, S. Alizon, Optimal COVID-19 epidemic control until vaccine deployment, medRxiv, (2020). Available from: https://www.medrxiv.org/content/10.1101/2020.04.02.20049189v2. |
| [10] | R. Engbert, M. M. Rabe, R. Kliegl, S. Reich, Sequential data assimilation of the stochastic SEIR epidemic model for regional COVID-19 dynamics, medRxiv, (2020). Available from: https://www.medrxiv.org/content/early/2020/06/30/2020.04.13.20063768.full.pdf. |
| [11] | S. E. Eikenberry, M. Mancuso, E. Iboi, T. Phan, K. Eikenberry, Y. Kuang, et al., To mask or not to mask: Modeling the potential for face mask use by the general public to curtail the COVID-19 pandemic, Infect. Dis. Model., 5 (2020), 293-308. |
| [12] |
S. W. Park, D. M. Cornforth, J. Dushoff, J. S. Weitz, The time scale of asymptomatic transmission affects estimates of epidemic potential in the COVID-19 outbreak, Epidemics, 31 (2020), 100392. doi: 10.1016/j.epidem.2020.100392
|
| [13] |
Y. Bai, L. Yao, T. Wei, F. Tian, D. Y. Jin, L. Chen, et al., Presumed asymptomatic carrier transmission of COVID-19, JAMA, 323 (2020), 1406-1407. doi: 10.1001/jama.2020.2565
|
| [14] | W. C. Roda, M. B. Varughese, D. Han, M. Y. Li, Why is it difficult to accurately predict the COVID-19 epidemic? Infect. Dis. Model., (2020). |
| [15] | G. Massonis, J. R. Banga, A. F. Villaverde, Structural identifiability and observability of compartmental models of the COVID-19 pandemic, (2020). Available on arXiv: arXiv: 2006.14295. |
| [16] |
I. Holmdahl, C. Buckee, Wrong but useful - what COVID-19 epidemiologic models can and cannot tell us, N. Engl. J. Med., 383 (2020), 303-305. doi: 10.1056/NEJMp2016822
|
| [17] |
F. Petropoulos, S. Makridakis, Forecasting the novel coronavirus COVID-19, PLoS ONE, 15 (2020), e0231236. doi: 10.1371/journal.pone.0231236
|
| [18] | L. Wynants, B. Van Calster, G. S. Collins, Prediction models for diagnosis and prognosis of COVID-19: systematic review and critical appraisal, BMJ, 369 (2020), m1328. |
| [19] |
Z. Wang, M. A. Andrews, Z. X. Wu, L. Wang, C. T. Bauch, Coupled disease-behavior dynamics on complex networks: a review, Phys. Life Rev., 15 (2015), 1-29. doi: 10.1016/j.plrev.2015.07.006
|
| [20] |
J. M. Epstein, J. Parker, D. Cummings, R. A. Hammond, Coupled contagion dynamics of fear and disease: Mathematical and computational explorations, PLoS ONE, 3 (2008), e3955. doi: 10.1371/journal.pone.0003955
|
| [21] | N. Perra, D. Balcan, B. Gonçalves, A. Vespignani, Towards a characterization of behavior-disease models, PLOS ONE, 6 (2011), 1-15. |
| [22] | S. Y. Del Valle, S. M. Mniszewski, J. M. Hyman, Modeling the Impact of Behavior Changes on the Spread of Pandemic Influenza, in Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases (eds. P. Manfredi and A. D'Onofrio), Springer New York, New York, NY, (2013), 59-77. |
| [23] |
J. A. Cui, X. Tao, H. Zhu, An SIS infection model incorporating media coverage, Rocky Mt. J. Math., 38 (2008), 1323-1334. doi: 10.1216/RMJ-2008-38-5-1323
|
| [24] |
E. P. Fenichel, C. Castillo-Chavez, M. G. Ceddia, G. Chowell, P. A. Gonzalez Parra, G. J. Hickling, et al., Adaptive human behavior in epidemiological models, Proc. Natl. Acad. Sci. U.S.A., 108 (2011), 6306-6311. doi: 10.1073/pnas.1011250108
|
| [25] | E. P. Fenichel, X. Wang, The Mechanism and Phenomena of Adaptive Human Behavior During an Epidemic and the Role of Information, in Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases (eds. P. Manfredi and A. D'Onofrio), Springer New York, New York, NY, (2013), 153-168. |
| [26] | P. Poletti, B. Caprile, M. Ajelli, S. Merler, Uncoordinated Human Responses During Epidemic Outbreaks, in Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases (eds. P. Manfredi and A. D'Onofrio), Springer New York, New York, NY, (2013), 79-91. |
| [27] |
C. Sun, W. Yang, J. Arino, K. Khan, Effect of media-induced social distancing on disease transmission in a two patch setting, Math. Biosci., 230 (2011), 87-95. doi: 10.1016/j.mbs.2011.01.005
|
| [28] |
M. A. Herrera-Valdez, M. Cruz-Aponte, C. Castillo-Chavez, Multiple outbreaks for the same pandemic: Local transportation and social distancing explain the different "waves" of A-H1N1 pdm cases observed in México during 2009, Math. Biosci. Eng., 8 (2011), 21. doi: 10.3934/mbe.2011.8.21
|
| [29] | S. A. Pedro, F. T. Ndjomatchoua, P. Jentsch, J. M. Tcheunche, M. Anand, C. T. Bauch, Conditions for a second wave of COVID-19 due to interactions between disease dynamics and social processes, (2020). Available on MedRxiv: https://www.medrxiv.org/content/10.1101/2020.05.22.20110502v1. |
| [30] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. Roy. Soc. Lond. A, 115 (1927), 700-721. doi: 10.1098/rspa.1927.0118
|
| [31] | S. Roy, COVID-19 reinfection: Myth or truth?, S.N. Compr. Clin. Med., (2020), 1-4. |
| [32] | O. Diekmann, J. A. P. Heesterbeek, Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis and Interpretation, in Wiley Series in Mathematical and Computational Biology, Chichester, Wiley, (1990). |
| [33] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
| [34] |
J. M. Heffernan, R. J. Smith, L. M. Wahl, Perspectives on the basic reproductive ratio, J. R. Soc. Interface, 2 (2005), 281-293. doi: 10.1098/rsif.2005.0042
|
| [35] | A. Hill, The possible effects of the aggregation of the molecules of haemoglobin on its dissociation curves, J. Physiol., 40 (1910), iv-vii. |
| [36] |
E. Dong, H. Du, L. Gardner, An interactive web-based dashboard to track COVID-19 in real time, Lancet Infect. Dis., 20 (2020), 533-534. doi: 10.1016/S1473-3099(20)30120-1
|
| [37] | Google, COVID-19 Community Mobility Reports, available from: https://www.google.com/covid19/mobility/ (Accessed: 11 Aug, 2020). |
| [38] | Apple, Mobility Trends Reports, available from: https://www.apple.com/covid19/mobility (Accessed: 11 Aug, 2020). |
| [39] | S. H. Strogatz, Nonlinear Dynamics and Chaos: with Applications to Physics, Biology, Chemistry, and Engineering, 2nd edition, Westview Press, Boulder, CO, 2015. |
| [40] | E. J. Routh, A Treatise on the Stability of a Given State of Motion: Particularly Steady Motion, Macmillan, 1877. |
| [41] |
A. Hurwitz, Ueber die bedingungen, unter welchen eine gleichung nur wurzeln mit negativen reellen theilen besitzt, Math. Ann., 46 (1895), 273-284. doi: 10.1007/BF01446812
|
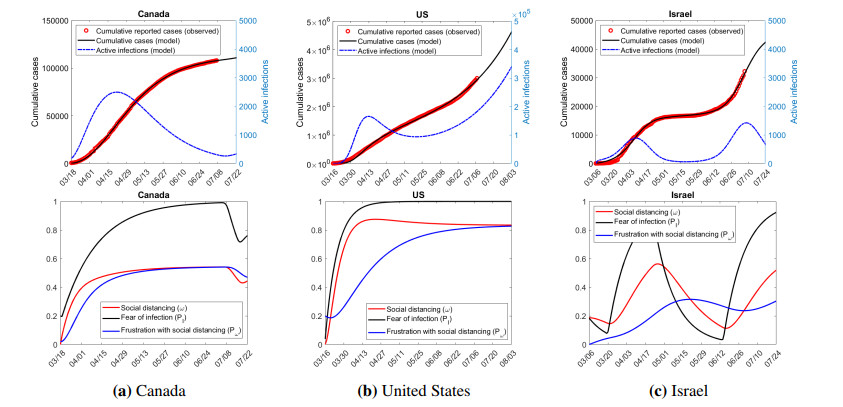
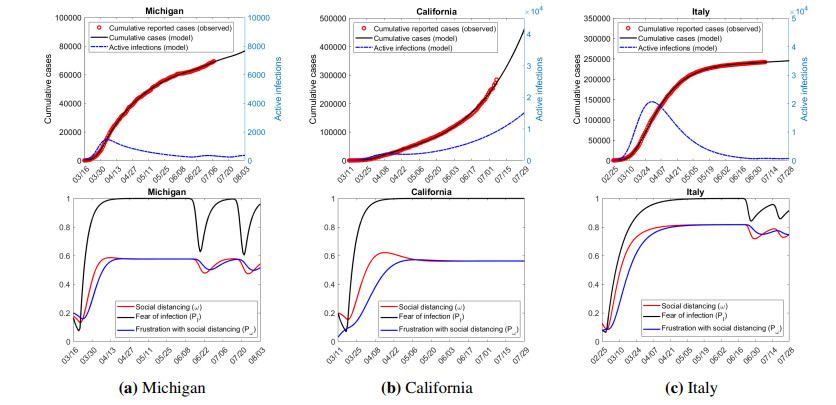
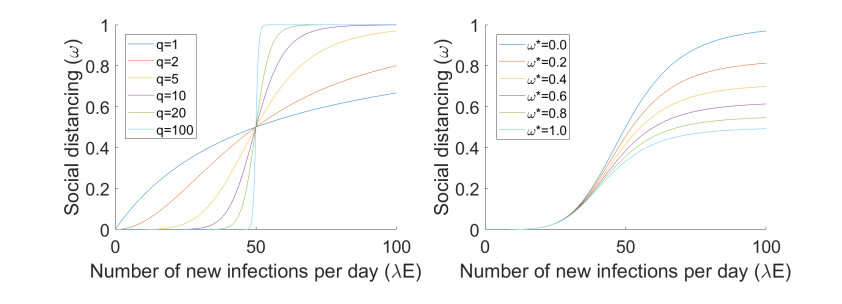
Figures(7) / Tables(3)

Matthew D. Johnston, Bruce Pell. A dynamical framework for modeling fear of infection and frustration with social distancing in COVID-19 spread[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7892-7915. doi: 10.3934/mbe.2020401







 DownLoad:
DownLoad: