Citation: Dario Madeo, Chiara Mocenni, Jean Carlo Moraes, Jorge P. Zubelli. The role of self-loops and link removal in evolutionary games on networks[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5287-5306. doi: 10.3934/mbe.2019264

| [1] | D. Madeo and C. Mocenni, Game Interactions and dynamics on networked populations, IEEE T. Automat. Contr., 60 (2015), 1801–1810. |
| [2] | A. Barrat, M. Barthelemy and A. Vespignani, Dynamical Processes on Complex Networks. Cambridge University Press, UK, 2008. |
| [3] | G. Ehrhardt, M. Marsili and F. Vega-Redondo, Diffusion and growth in an evolving network, Int. J. Game Theory, 334 (2006), 383–397. |
| [4] | V. Colizza, A. Barrat, M. Barthélemy, et al., The role of the airline transportation network in the prediction and predictability of global epidemics, P. Natl. Acad. Sci. USA, 103 (2006), 2015–2020. |
| [5] | V. Colizza and A. Vespignani, Epidemic modeling in metapopulation systems with heterogeneous coupling pattern: Theory and simulations, J. Theor. Biol., 251 (2008), 450–467. |
| [6] | S. Tully, M. G. Cojocaru and C. T. Bauch, Multiplayer games and HIV transmission via casual encounters, Math. Biosci. Eng., 14 (2017), 359–376. |
| [7] | M. D'Orsogna and M. Perc, Statistical physics of crime: A review, Phys. Life Rev., 12 (2015), 1–21. |
| [8] | D. Madeo, L. R. Comolli and C. Mocenni, Emergence of microbial networks as response to hostile environments, Front. Microbiol., 5 (2014), 407. |
| [9] | N. Quijano, C. Ocampo-Martinez, J. Barreiro-Gomez, et al., The role of population games and evolutionary dynamics in distributed control systems: The advantages of evolutionary game theory, IEEE Contr. Sys. Mag., 37 (2017), 70–97. |
| [10] | R. Gray, A. Franci, V. Srivastava, et al., Multi-agent decision-making dynamics inspired by honeybees, IEEE T. Contr. Netw. Sys., 5 (2018), 793–806. |
| [11] | F. C. Santos, J. M. Pacheco and T. Lenaerts, Evolutionary dynamics of social dilemmas in structured heterogeneous populations, P. Natl. Acad. Sci. USA, 103 (2006), 3490–3494. |
| [12] | H. Ohtsuki and M. A. Nowak, The replicator equation on graphs, J. Theor. Biol., 243 (2006), 86–97. |
| [13] | T. Konno, A condition for cooperation in a game on complex networks, J. Theor. Biol., 269 (2011), 224–233. |
| [14] | J. Gómez-Gardenes, I. Reinares, A. Arenas, et al., Evolution of cooperation in multiplex networks, Sci. Rep., 2 (2012), 620. |
| [15] | S. M. Cameron and A. Cintrón-Arias, Prisoner's Dilemma on real social networks: Revisited, Math. Biosci. Eng., 10 (2013), 1381–1398. |
| [16] | D. G. Rand, M. A. Nowak, J. H. Fowler, et al., Static network structure can stabilize human cooperation, P. Natl. Acad. Sci. USA, 11 (2014), 17093–17098. |
| [17] | B. Allen, G. Lippner, Y. Chen, et al., Evolutionary dynamics on any population structure, Nature, 544 (2017), 227. |
| [18] | B. Fotouhi, N. Momeni, B. Allen, et al., Evolution of Cooperation on Stochastic Block Models, preprint, arXiv:1807.03093. |
| [19] | J. Weibull, Evolutionary Game Theory, MIT Press, Cambridge, MA, 1995. |
| [20] | J. Hofbauer and K. Sigmund, Evolutionary game dynamics, B. Am. Math. Soc, 40 (2003) 479–519. |
| [21] | M. A. Nowak, Evolutionary Dynamics: Exploring the Equations of Life, Belknap Press of Harvard University Press, Harvard, MA, 2006. |
| [22] | G. Iacobelli, D. Madeo and C. Mocenni, Lumping evolutionary game dynamics on networks, J. Theor. Biol., 407 (2016), 328–338. |
| [23] | D. Pais, C. H. Caicedo-Nùñez and N. E. Leonard, Hopf bifurcations and limit cycles in evolutionary network dynamics, SIAM J. Appl. Dyn. Syst., 11 (2012), 1754–1884. |
| [24] | W. Ren and R. Beard, Consensus seeking in multiagent systems under dynamically changing interaction topologies, IEEE T. Automat. Contr., 50 (2005), 655–661. |
| [25] | R. Olfati-Saber, A. Fax and R. Murray, Consensus and cooperation in networked multi-agent systems, P. IEEE, 95 (2007), 215–233. |
| [26] | B. Kozma and A. Barrat, Consensus formation on adaptive networks, Phys. Rev. E, 77 (2008), 016102. |
| [27] | G. Punzo, G. F. Young, M Macdonald, et al., Using network dynamical influence to drive consensus, Sci. Rep., 6 (2016), 26318. |
| [28] | A. Traulsen, F. C. Santos and J. M. Pacheco, Evolutionary Games in Self-Organizing Populations, in Adaptive networks: Theory, Models and Applications (eds. T. Gross and H. Sayama), Springer Berlin Heidelberg, Germany, (2009), 253–267. |
| [29] | S. Boccaletti, V. Latora, Y. Moreno, et al., Complex networks: Structure and dynamics, Phys. Rep., 424 (2006), 175–308. |
| [30] | Y. Bramoullé and R. Kranton, Games Played on Networks, in The Oxford Handbook of the Economics of Networks (eds. Y. Bramoullé, A. Galeotti and B. Rogers), Oxford University Press. Available from: http://www.oxfordhandbooks.com/view/10.1093/oxfordhb/9780199948277.001. 0001/oxfordhb-9780199948277. |
| [31] | A. Banerjee, A. G. Chandrasekhar, E. Duflo, et al., Gossip: Identifying Central Individuals in a Social Network, preprint, arXiv:1406.2293v3. |
| [32] | D. Madeo and C. Mocenni, Self-regulation promotes cooperation in social networks, preprint, arXiv:1807.07848. |
| [33] | M. Newman, Network: An introduction, Oxford University Press, 2010. |
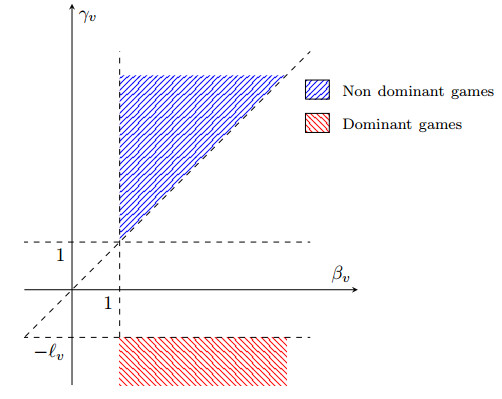
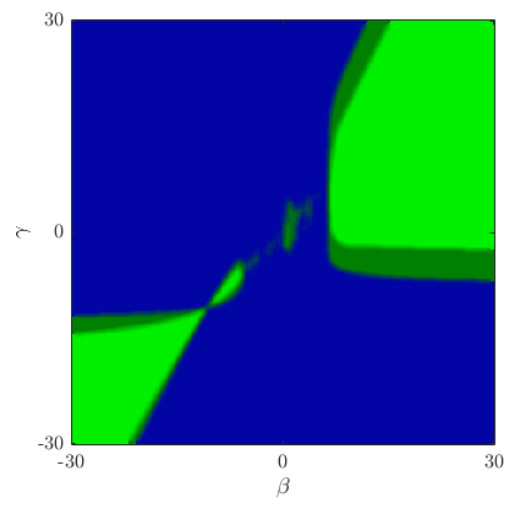
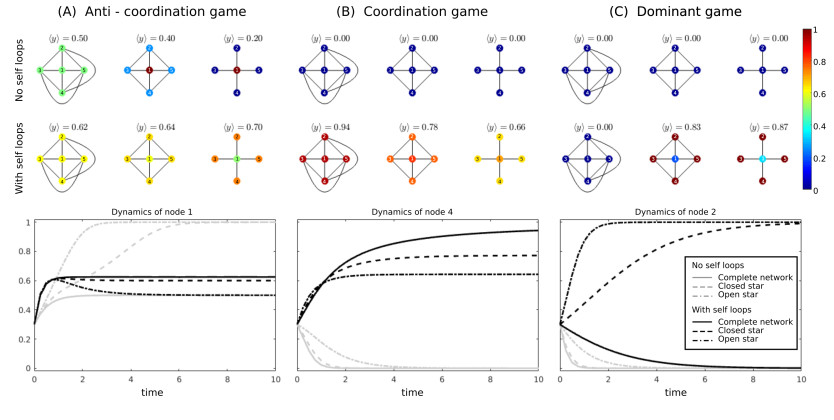
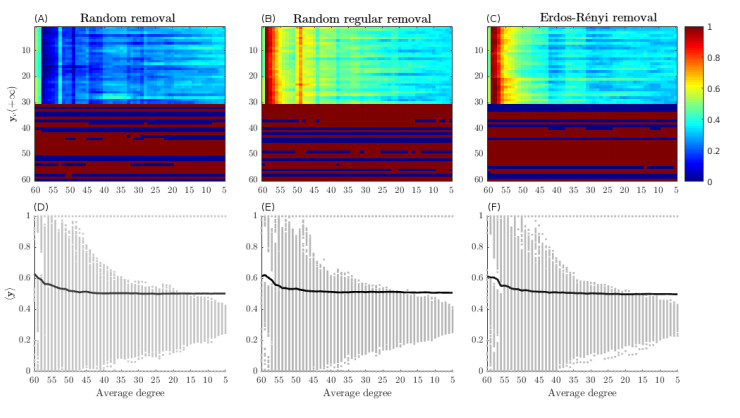
Figures(5)

Dario Madeo, Chiara Mocenni, Jean Carlo Moraes, Jorge P. Zubelli. The role of self-loops and link removal in evolutionary games on networks[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5287-5306. doi: 10.3934/mbe.2019264







 DownLoad:
DownLoad: