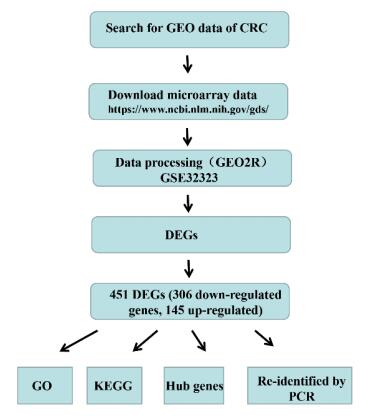
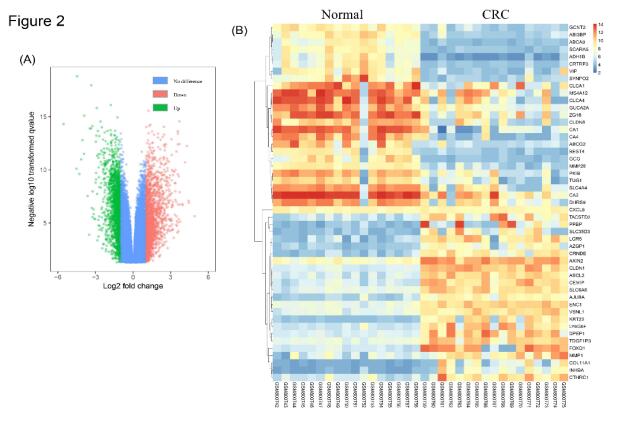
Citation: Zhongwei Zhao, Xiaoxi Fan, Lili Yang, Jingjing Song, Shiji Fang, Jianfei Tu, Minjiang Chen, Liyun Zheng, Fazong Wu, Dengke Zhang, Xihui Ying, Jiansong Ji. The identification of a common different gene expression signature in patients with colorectal cancer[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2942-2958. doi: 10.3934/mbe.2019145

| [1] | K. Yang, F. Zhang, P. Han, et al., Metabolomics approach for predicting response to neoadjuvant chemotherapy for colorectal cancer, Metabolomics, 9 (2018), 110–121. |
| [2] | R. L. Siegel, K. D. Miller, S. A. Fedewa, et al., Colorectal cancer statistics, 2017. CA Cancer J. Clin., 3 (2017), 104–117. |
| [3] | P. M. Yang, Y. T. Li, C. T. Shun, et al., Zebularine inhibits tumorigenesis and stemness of colorectal cancer via p53-dependent endoplasmic reticulum stress, Sci. Rep., 11 (2013), 3219.4. S. A. Noonan, M. E. Morrissey, P. Martin, et al., Tumour vasculature immaturity, oxidative damage and systemic inflammation stratify survival of colorectal cancer patients on bevacizumab treatment, Oncotarget, 12 (2018), 10536–10548. |
| [4] | 5. Y. Liu, G. Wang, Y. Yang, et al., Increased TEADexpression and nuclear localization in colorectal cancer promote epithelial-mesenchymal transition and metastasis in a YAP-independent manner, Oncogene, 21 (2016), 2789–2795. |
| [5] | 6. A. G. Long, E. T. Lundsmith and K. E. Hamilton, Inflammation and colorectal cancer, Curr. Colorectal Cancer Rep., 4 (2017), 341–3. |
| [6] | 7. H. Zheng, L. Yu, S. Luo, et al., miR-29inhibits the metastasis and epithelial-mesenchymal transition of colorectal cancer by targeting S100A4, BMC Cancer, 1 (2017), 140–147. |
| [7] | 8. A. Mahasneh, F. Alshaheri and E. Jamal, Molecular biomarkers for an early diagnosis, effective treatment and prognosis of colorectal cancer: Current updates, Exp. Mol. Pathol., 3 (201, 475–483. |
| [8] | 9. N. Li, L. Li and Y. Chen, The identification of core gene expression signature in hepatocellular carcinoma, Oxid. Med. Cell Longev., 4 (201, 1–15. |
| [9] | 10. L. Ling, L. Ning, C. He, et al., Proteomic analysis of differentially expressed proteins in kidneys of brain dead rabbits, Mol. Med. Rep., 1 (2017), 215–223. |
| [10] | 11. T. Braunschweig, J. Y. Chung, S. M. Hewitt, Tissue microarrays: bridging the gap between research and the clinic, Expert Rev. Proteom., 3 (2005), 325–336. |
| [11] | 12. A. Khamas, T. Ishikawa, K. Shimokawa, et al., Screening for epigenetically masked genes in colorectal cancer Using 5-Aza-2'-deoxycytidine, microarray and gene expression profile. Cancer Genom. Proteom., 2 (2012), 67–7513. G. Dennis, B. T. Sherman, D. A. Hosack, et al., DAVID: Database for Annotation, visualization, and Integrated Discovery, Genome Biol., 5 (2003), 3. |
| [12] | 14. Z. Tang, C. Li, B. Kang, et al., GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses, Nucleic Acids Res., 1 (2017), 98–102. |
| [13] | 15. L. P. Sun, Y. Guo, Y. X. Zhang, et al., Antioxidant and anti-tyrosinase activities of phenolic extracts from rape bee pollen and inhibitory melanogenesis by cAMP/MITF/TYR pathway in B16 mouse melanoma cells, Front. Pharmaco., 176 (2017), 46–51. |
| [14] | 16. A. Ikin, C. Riveros, P. Moscato, et al., The Gene Interaction Miner: a new tool for data mining contextual information for protein–protein interaction analysis, Bioinformatics, 2 (2010), 283–284. |
| [15] | 17. M. Arnold, M. S. Sierra, M. Laversanne, et al., Global patterns and trends in colorectal cancer incidence and mortality, Gut, 4 (2017), 683–691. |
| [16] | 18. D. K. Rex, C. R. Boland, J. A. Dominitz, et al., Colorectal cancer screening: recommendations for physicians and patients from the U.S. multi-society task force on colorectal cancer, Gastroenterology, 7 (2017), 307–323. |
| [17] | 19. E. V. Cutsem, A. Cervantes, R. Adam, et al., ESMO consensus guidelines for the management of patients with metastatic colorectal cancer, Ann. Oncol., 8 (2016), 1386–1422. |
| [18] | 20. V. P. Deenadayalu and D. K. Rex, Colorectal cancer screening: a guide to the guidelines, Rev. Gasteroenterol. Di., 7 (2016), 204. |
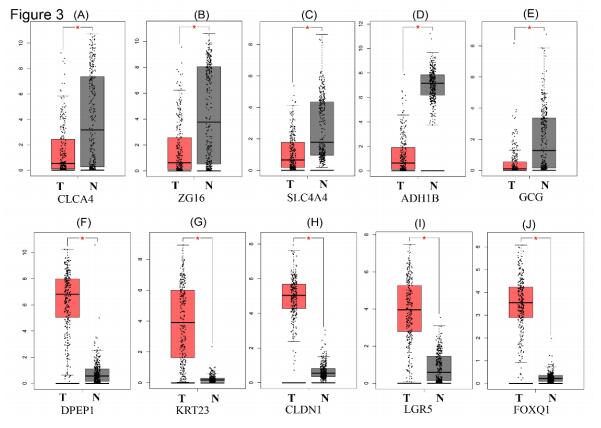
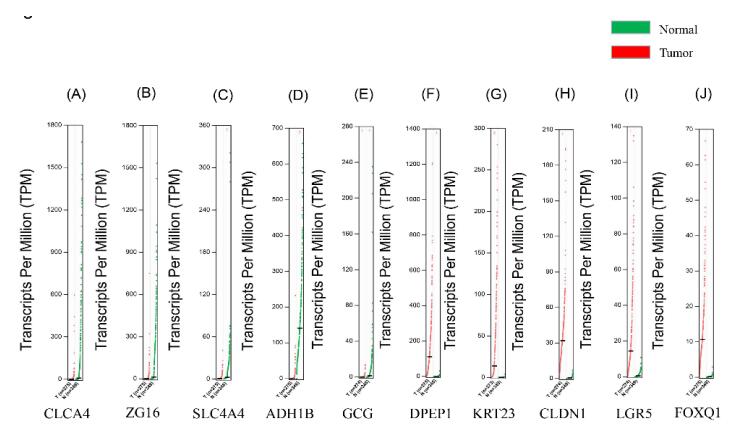
| [19] | 21. M. Hui, W. Li, L. A. Boardman, et al., Loss of ZG16 is associated with molecular and clinicopathological phenotypes of colorectal cancer, BMC Cancer, 1 (2018), 433–441. |
| [20] | 22. P. A. Eisenach, E. Soeth, C. Röder, et al., Dipeptidase 1 (DPEP1) is a marker for the transition from low-grade to high-grade intraepithelial neoplasia and an adverse prognostic factor in colorectal cancer, Brit. J. Cancer., 3 (3), 694–703. |
| [21] | 23. S. Pyronnet, C. Bousquet, S. Najib, et al., Antitumor effects of somatostatin, Mol. Cell Endocrinol., 1 (2008), 230–237. |
| [22] | 24. K. Leiszter, F. Sipos, O. Galamb, et al., Promoter Hypermethylation-Related Reduced Somatostatin Production Promotes Uncontrolled Cell Proliferation in Colorectal Cancer, Plos One, 2 (2015), e0118332. |
| [23] | 25. Y. Mori, K. Cai, Y. Cheng, et al., A genome-wide search identifies epigenetic silencing of somatostatin, tachykinin-1, and 5 other genes in colon cancer, Gastroenterology, 3 (2006), 797–808. |
| [24] | 26. Y. Liu, H. C. Min, C. K. Tham, et al., Methylation of serum SST gene is an independent prognostic marker in colorectal cancer, Am. J. Cancer Res., 9 (2016), 2098–3008. |
| [25] | 27. N. Nagarsheth, M. S. Wicha and W. Zou, Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy, Nat. Rev. Immunol., 9 (2017), 559–572. |
| [26] | 28. H. Verbeke, S. Struyf, G. Laureys, et al., The expression and role of CXC chemokines in colorectal cancer, Cytokine Growth F. R., 5 (2011), 345–358. |
| [27] | 29. O. Oladipo, S. Conlon, A. O'Grady, et al., The expression and prognostic impact of CXC-chemokines in stage II and III colorectal cancer epithelial and stromal tissue, Br. J. Cancer, 3 (2011),480–487. |
| [28] | 30. K. Rupertus, J. Sinistra, C. Scheuer, et al., Interaction of the chemokines I-TAC (CXCL11) and SDF-1 (CXCL12) in the regulation of tumor angiogenesis of colorectal cancer, Clin. Exp. Metastasis, 4 (2014), 447–459. |
| [29] | 31. X. Shen, A. Artinyan, D. Jackson, et al., Chemokine receptor CXCR4 enhances proliferation in pancreatic cancer cells through AKT and ERK dependent pathways, Pancreas, 1(2010), 81–87. |
| [30] | 32. Y. Fang, F. C. Henderson, Q. Yi, et al., Chemokine CXCL16 expression suppresses migration and invasiveness and induces apoptosis in breast cancer cells, Mediat. Inflamm., 8 (2014), 478641. |
| [31] | 33. J. Ehling and F. Tacke, Role of chemokine pathways in hepatobiliary cancer, Cancer Lett., 2 (2016), 173–183. |
Figures(8) / Tables(4)

Zhongwei Zhao, Xiaoxi Fan, Lili Yang, Jingjing Song, Shiji Fang, Jianfei Tu, Minjiang Chen, Liyun Zheng, Fazong Wu, Dengke Zhang, Xihui Ying, Jiansong Ji. The identification of a common different gene expression signature in patients with colorectal cancer[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2942-2958. doi: 10.3934/mbe.2019145







 DownLoad:
DownLoad: