Citation: Yukihiko Nakata, Ryosuke Omori. The change of susceptibility following infection can induce failure to predict outbreak potential by $\mathcal{R}_{0}$[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 813-830. doi: 10.3934/mbe.2019038

| [1] | R.M. Anderson and R.M. May, Infectious diseases of humans: dynamics and control, Vol. 28. Oxford: Oxford University Press, 1992. |
| [2] | J. Arino, C. C. McCluskey and P. van den Driessche, Global results for an epidemic model with vaccination that exhibits backward bifurcation, SIAM J. Appl. Math., 64 (2003), 260–276. |
| [3] | N. Arinaminpathy, J. S. Lavine and B. T. Grenfell, Self-boosting vaccines and their implications for herd immunity, PNAS, 109 (2012), 20154-20159. |
| [4] | M.V. Barbarossa, A. Denes, G. Kiss, Y. Nakata and G. R´'ost, Zs. Vizi, Transmission dynamics and final epidemic size of Ebola Virus Disease outbreaks with varying interventions, PLoS One, 10 (2015), e0131398. |
| [5] | W. Dejnirattisai, A. Jumnainsong, N. Onsirisakul, P. Fitton, S. Vasanawathana, W. Limpitikul, C. Puttikhunt, C. Edwards, T, Duangchinda , S. Supasa, K. Chawansuntati, P. Malasit, J. Mongkolsapaya and G. Screaton, Cross-reacting antibodies enhance dengue virus infection in humans, Science, 328 (2010), 745-748. |
| [6] | F. Brauer, Backward bifurcations in simple vaccination models, J. Math. Anal. Appl., 29 (2004), 418–431. |
| [7] | O. Diekmann, J. A. P. Heesterbeek and J. A. J. Metz, On the definition and the computation of the basic reproduction ratio $\mathcal{R}_{0}$ in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. |
| [8] | O. Diekmann, J. A. P. Heesterbeek and J. A. J. Metz, The legacy of Kermack and McKendrick, Epidemic Models: Their Structure and Relation to Data, (D. Mollison, ed.) (1995), 95–115. |
| [9] | O. Diekmann and R. Montijn, Prelude to Hopf bifurcation in an epidemic model: analysis of a characteristic equation associated with a nonlinear Volterra integral equation, J. Math. Biol., 14 (1982), 117–127. |
| [10] | O. Diekmann, H. Heesterbeek and T. Britton, Mathematical tools for understanding infectious disease dynamics, Princeton University Press, 2012. |
| [11] | W.J. Edmunds, O.G. Van de Heijden, M. Eerola and N. J. Gay, Modelling rubella in Europe, Epidemiol. Infect., 125 (2000), 617–634. |
| [12] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [13] | M.G.M. Gomes, L.J. White and G.F. Medley, Infection, reinfection, and vaccination under suboptimal immune protection: epidemiological perspectives, J. Theor. Biol., 228 (2004), 539–549. |
| [14] | D. Greenhalgh, O. Diekmann and M. C. M de Jong, Subcritical endemic steady states in mathematical models for animal infections with incomplete immunity, Math. Biosci., 165 (2000), 1–25. |
| [15] | C. Hall, E. Walsh, C. Long and K. Schnabel, Immunity to and frequency of reinfection with respiratory syncytial virus, J. Infect. Dis., 163(1991), 693–698. |
| [16] | H.W. Hethcote, H.W. Stech and P. van den Driessche, Nonlinear oscillations in epidemic models, SIAM J. Appl. Math., 40 (1981), 1–9. |
| [17] | H. Inaba, Kermack and McKendrick revisited: the variable susceptibility model for infectious diseases, J. Ind. Appl. Math., 18 (2001), 273–292. |
| [18] | H. Inaba, Endemic threshold analysis for the Kermack-McKendrick reinfection model, Josai Math. Mono., 9 (2016), 105–133. |
| [19] | H. Inaba, Age-structured population dynamics in demography and epidemiology, Springer, 2017. |
| [20] | D. Isaacs, D. Flowers, J. R. Clarke, H. B. Valman and M. R. MacNaughton, Epidemiology of coronavirus respiratory infections, Arch. Dis. Child., 58 (1983), 500–503. |
| [21] | G. Katriel, Epidemics with partial immunity to reinfection, Math. Biosci., 228 (2010), 153–159. |
| [22] | W.O. Kermack and A.G. McKendrick, Contributions to the mathematical theory of epidemics-II. The problem of endemicity. Proceedings of the Royal Society, 138A (1932), 55–83; Reprinted in Bull, Math. Biol., 53 (1991), 57–87. |
| [23] | W.O. Kermack and A.G. McKendrick, Contributions to the mathematical theory of epidemics-III. Further studies of the problem of endemicity. Proceedings of the Royal Society, 141A (1933), 94–122; Reprinted in Bull, Math. Biol., 53 (1991), 89–118. |
| [24] | R. Kohlmann, A. Salmen and A. Chan, Serological evidence of increased susceptibility to varicella-zoster virus reactivation or reinfection in natalizumab-treated patients with multiple sclerosis, Mult. Scler. J., 21 (2015), 1823–1832. |
| [25] | J.R. Kremer, F. Schneider and C.P. Muller, Waning antibodies in measles and rubella vaccinees-a longitudinal study, Vaccine, 24 (2006), 2594–2601. |
| [26] | C.M. Kribs-Zaleta and J.X. Velasco-Hernandez, A simple vaccination model with multiple endemic states, Math. Biosci., 164 (2000), 183–201. |
| [27] | J.S. Lavine, A. A. King and O. N. Bjørnstad, Natural immune boosting in pertussis dynamics and the potential for long-term vaccine failure, PNAS, 108 (2011), 7259–7264. |
| [28] | Y. Nakata, Y. Enatsu, H. Inaba, T. Kuniya, Y. Muroya and Y. Takeuchi, Stability of epidemic models with waning immunity, SUT J. Math., 50 (2014), 205–245. |
| [29] | C.A. Siegrist, Vaccine immunology, Vaccines, 5 (2008), 1725. |
| [30] | T.J.W. van de Laar, R. Molenkamp, C. V. D. Berg, Frequent HCV reinfection and superinfection in a cohort of injecting drug users in Amsterdam, J. Hepatol., 51 (2009), 667–674. |
| [31] | S. Verver, Rate of reinfection tuberculosis after successful treatment is higher than rate of new tuberculosis, Am. J. Resp. Crit. Care, 171 (2005), 1430–1435. |
| [32] | G. Wei, H. Bull, X. Zhou and H. Tabel, Intradermal infections of mice by low numbers of African trypanosomes are controlled by innate resistance but enhance susceptibility to reinfection, J.Infect. Dis., 203 (2011), 418–429. |
| [33] | S.S. Whitehead, J.E. Blaney, A.P. Durbin and B.R. Murphy, Prospects for a dengue virus vaccine, Nat. Rev. Microbiol., 5 (2007), 518–528. |
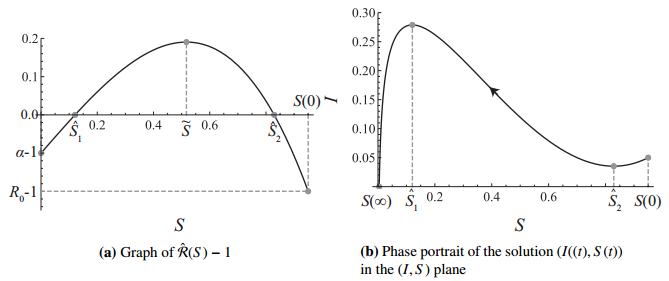
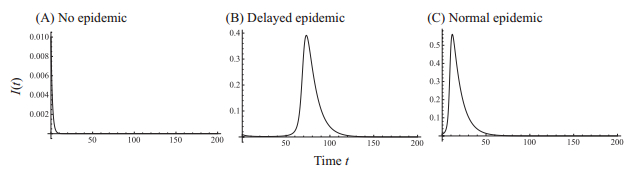
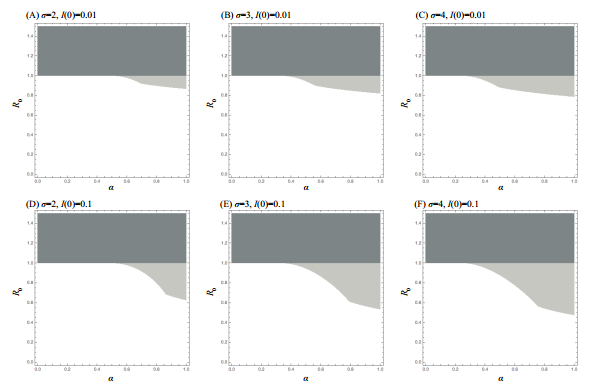
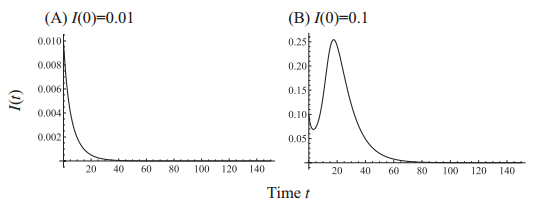
Figures(7)

Yukihiko Nakata, Ryosuke Omori. The change of susceptibility following infection can induce failure to predict outbreak potential by $\mathcal{R}_{0}$[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 813-830. doi: 10.3934/mbe.2019038







 DownLoad:
DownLoad: